Для лучшего понимания темы рекомендую прочесть предыдущую публикацию
Далее привожу еще одну главу из книги Николая Левашова "Неоднородная вселенная":
"
Глава 4. Необходимые и достаточные условия
возникновения жизни во Вселенной
4.1. Постановка вопроса
Вопрос о возникновении жизни на нашей планете всегда был «камнем преткновения». С древних времён философы, учёные пытались разгадать тайну жизни. Создавались разные теории, гипотезы о природе живой материи. Все они базируются на постулатах (понятиях, принимаемых без доказательств). Чтобы сохранить эти теории жизнеспособными, позднее вводились новые и новые постулаты. В настоящее время все существующие научные теории имеют в своём фундаменте десятки, а порой и сотни постулатов. К их числу относится и современная физика. Информация, которую человечество накопило к концу двадцатого века, полностью делает эти теории несостоятельными. Явления, которые учёные наблюдают, посредством приборов или визуально, есть проявления реальных законов природы. Но, реальные законы природы формируются на уровнях макрокосмоса и микрокосмоса. Всё, с чем человек соприкасается в своей жизни, находится между макрокосмосом и микрокосмосом. Именно поэтому, когда человек с помощью приборов смог заглянуть в микромир, он впервые столкнулся с законами природы, а не с их проявлениями. Материя не появилась из неоткуда. Всё гораздо проще и сложнее одновременно: то, что человек знает о материи и думает, как о завершённом, абсолютном понятии, на самом деле, является лишь маленькой частью этого понятия. Материя действительно никуда не исчезает и ниоткуда не появляется; действительно существует Закон Сохранения Материи, только он не такой, каким его представляют люди. Таким образом, существующие научные теории, основанные на постулатах, оказались мёртворождёнными. Они не смогли дать какого-либо стройного и логического объяснения. Невозможность существующих теорий объяснить условия и причины зарождения жизни не извиняют это незнание. Жизнь на нашей планете появилась более четырёх миллиардов лет назад и её развитие привело к появлению разумности, но существующая цивилизация до сих пор, не может ответить на простой вопрос: что такое жизнь, как она возникла из так называемой неживой материи? Каким образом и почему, неживая материя, вдруг, преобразуется в живую? Без понимания этого вопроса, человечество не может называть себя разумной расой, а только неразумным малышом, для которого пришла пора набираться ума-разума. Итак, какие условия должны были возникнуть на планете, при которых возможно зарождение жизни?
4.2. Условия зарождения жизни на планетах
Прежде, чем объяснить природу зарождения жизни, в первую очередь необходимо определить, какие условия должны существовать, чтобы на планете могла зародиться, по крайней мере, белковая жизнь. Девять планет солнечной системы — наглядный пример этому. В данный момент, только на планете Земля существуют необходимые и достаточные условия для жизни или, по крайней мере, сложноорганизованной живой материи. И первоочередной задачей является определение этих условий. Исходя из понимания вышеупомянутых процессов, происходящих на макро- и микроуровнях пространства, можно выделить следующие условия, необходимые для зарождения жизни:
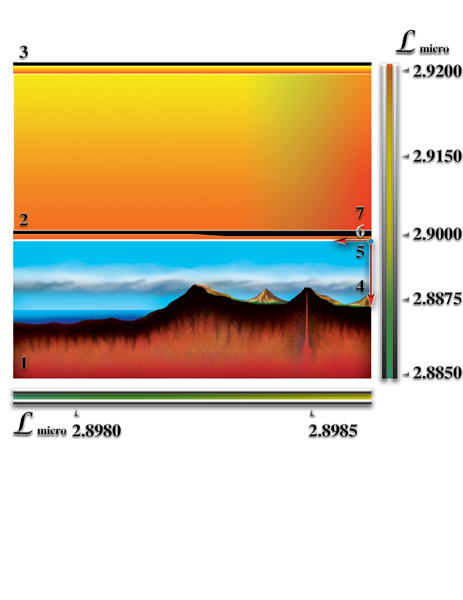
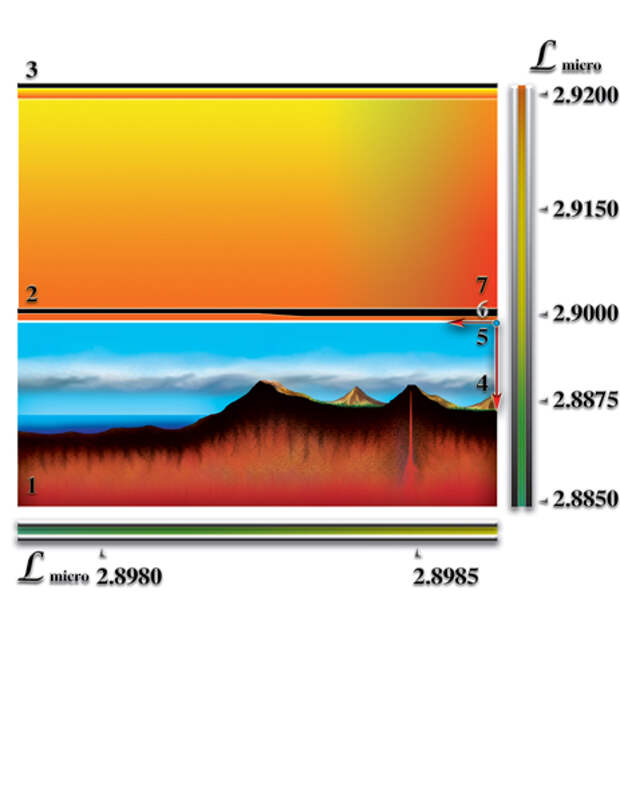
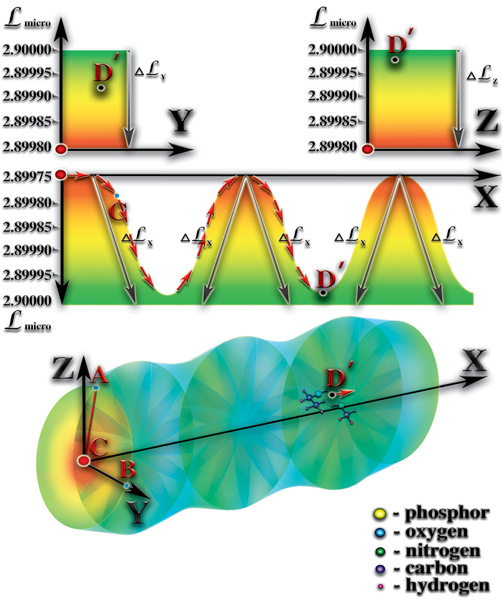
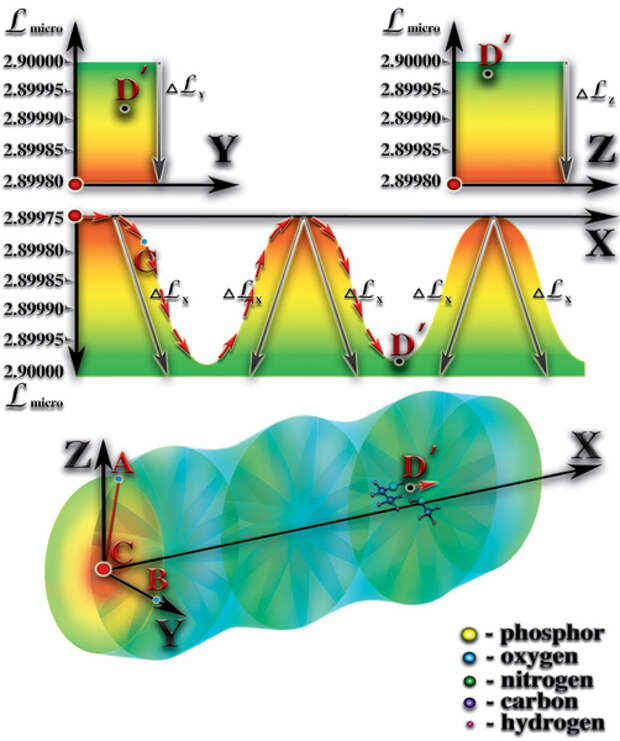
1. Наличие постоянного перепада мерности ς. Величина постоянного перепада мерности и коэффициент квантования пространства γi (определяющий количество форм материй данного типа, которые могут слиться в пределах этого перепада) определяют эволюционный потенциал возможной жизни. Кратность этих величин — критерий, дающий представление о количестве качественных барьеров (уровней), возникающих внутри этого перепада мерности. Количество барьеров характеризует качественное многообразие возможной жизни. В том числе, возможность появления разума и его развития. Мерность макропространства, после завершения формирования планеты, возвращается к исходному уровню, который был до взрыва сверхновой звезды. После завершения процесса образования возникает постоянный перепад мерности между уровнем мерности физически плотного вещества (2.89915) и уровнем мерности окружающего макрокосмоса (3.00017). Таким образом, постоянный перепад мерности является необходимым условием возникновения жизни. Важное значение имеет величина этого перепада. Именно величина перепада определяет эволюционный потенциал живой материи, жизни. Минимальный перепад мерности, при котором возможно зарождение жизни, должен быть равен:
ς = 1 γi (ΔL) (4.2.1)
Появление элементов разума и зарождение памяти, без которой невозможно развития разума, возможно при перепаде мерности, равном:
ς = 2 γi (ΔL) (4.2.2)
Необходимым условием для возникновения разума и его эволюции является перепад мерности:
ς = 3 γi (ΔL) (4.2.3)
Таким образом, используя перепад мерности, как критерий, можно говорить о требовании к качественной структуре пространства-вселенной (для нашего пространства-вселенной (γi (ΔL) = 0.020203236...). Только пространства-вселенные, образованные тремя и большим количеством форм материй имеют необходимые условия для зарождения жизни и разума
2. Наличие воды. Вода является основой органической жизни на нашей планете. Конечно же, существуют формы жизни не только на белковой основе. Но для начала, необходимо проследить закономерности возникновения белковой жизни. Необходимо понять, что происходит в нашем собственном доме перед тем, как заглядывать в чужие.
3. Наличие атмосферы. Атмосфера является наиболее динамичной, активной частью планеты. Она быстро и резко реагирует на изменения состояния внешней среды, что очень важно для возникновения жизни. Наличие в атмосфере кислорода и углекислого газов — знак наличия на планете белковой жизни. Атмосфера не должна быть очень плотной и чрезмерно разрежённой. При очень плотной атмосфере излучения звезды не достигают поверхности планеты и не нагревают её. При этом нижние слои атмосферы не поглощают излучения звезды и тепловые излучения поверхностных слоёв планеты. В результате, перепад мерности между освещённой и ночной частями поверхности планеты не возникает. И, как следствие, не возникает движение атмосферных масс в нижних слоях атмосферы. При отсутствии градиента (перепада) мерности вдоль поверхности планеты, не возникают атмосферные электрические разряды. В чрезмерно разрежённой атмосфере нижние слои имеют возможность поглощать излучения звезды и тепловые излучения поверхности. Но, при этом, не возникает движение атмосферных масс, как результат её чрезмерной разрежённости. Как известно, величина и плотность атмосферы определяется размером и массой планеты. Поэтому, только планеты, соизмеримые по размерам и массе с нашей планетой Землёй имеют максимально благоприятные условия для возникновения белковой жизни. Атмосфера не должна быть ни чрезмерно «тяжёлой», ни чрезмерно «лёгкой».
4. Наличие периодической смены дня и ночи. Планетарные сутки не должны быть очень короткими или очень длинными. Планеты с продолжительностью планетарных суток в пределах диапазона 18-48 земных часов имеют максимально благоприятные условия для возникновения жизни. При массовом поглощении фотонов света атомами поверхностного слоя больших площадей, происходит увеличение уровня мерности этого слоя на некоторую величину ΔL. Эта величина соответствует амплитуде волн, которые поглощаются поверхностным слоем планеты (инфракрасное, оптическое, ультрафиолетовое излучения Солнца). В результате этого, перепад между уровнями мерности атмосферы и поверхности планеты в зоне поглощения уменьшается на величину ΔL, в то время, как неосвещённая или ночная часть поверхности сохраняет прежний перепад уровней мерности между атмосферой и поверхностью. Таким образом, возникает перепад мерности между освещённой и неосвещённой зонами поверхности планеты. Возникает параллельный поверхности планеты перепад (градиент) мерности. Определяющее значение имеет величина этого перепада. Дело в том, что молекулы атмосферы находятся под воздействием гравитационного поля планеты, существующего постоянно, как следствие формирования в зоне неоднородности макропространства постоянного перепада мерности, направленного от внешних границ к центру зоны неоднородности.
Гравитационное поле планеты компенсируется тем, что каждый атом или молекула атмосферы имеют уровни собственной мерности, очень близкие к верхней границе диапазона устойчивости физически плотного вещества. Вступает в силу, так называемый, «эффект поплавка», когда каждая молекула или атом стремятся к положению максимально устойчивого состояния равновесия. Именно, благодаря этому, молекулы и атомы атмосферы не падают на поверхность планеты, как молекулы и атомы более тяжёлых элементов. Перепад (градиент) мерности между дневной и ночной зонами направлен вдоль поверхности планеты, что приводит в движение свободные материи параллельно её поверхности от зоны с большим уровнем мерности (освещённая поверхность) к зоне с меньшим уровнем мерности (неосвещённая поверхность). В результате появления второго направления движения свободных материй параллельно поверхности, возникает перепад атмосферного давления (Рис. 4.2.1)
{kind=link}
и уменьшается сила тяжести.
Так как молекулы атмосферы не связаны между собой в жёсткие (твёрдое состояние вещества) или полужёсткие системы (жидкое состояние вещества), то перепад мерности пространства вдоль поверхности приводит к тому, что поток свободных материй увлекает за собой молекулы, образующие атмосферу. Воздушные массы приходят в движение, возникает ветер. При этом, «разогретые» молекулы (молекулы, поглотившие солнечные излучения) перемещаются на неосвещённую территорию, где происходит спонтанное (самопроизвольное) излучение ими волн. Другими словами, вследствие того, что собственный уровень мерности этих молекул выше собственного уровня атмосферы неосвещённой поверхности, этот перепад, между мерностью среды и собственной мерностью разогретых молекул, вызывает неустойчивое состояние последних и провоцирует спонтанное излучение молекулами волн. «Холодные» молекулы, в свою очередь, имеют уровень собственный мерности ниже собственного уровня мерности освещённой территории, что провоцирует массовое поглощение излучений Солнца и тепловых излучений освещённой поверхности. Постепенно происходит выравнивание между собственным уровнем мерности освещенной поверхности и собственным уровнем мерности молекул. При этом, если собственный уровень мерности «холодных» молекул значительно отличается от собственного уровня мерности освещённой территории, происходит снижение последнего. Когда собственный уровень мерности освещённой территории опускается до уровня, так называемой, точки «росы», молекулы воды из газообразного состояния переходят в жидкое. Выпадает роса. Если это происходит на уровне облачности, процесс каплеобразования приобретает цепной характер, и выпадает дождь. При этом, состояние качественного барьера между вторым и физическим уровнями возвращается к норме. В случае, когда этот процесс происходит быстро и резко, скопившиеся на уровне качественного барьера свободные материи стекают лавинообразно. И, как следствие, возникают атмосферные электрические разряды — молнии. Аналогией этому процессу может послужить плотина на реке, у которой открыли все шлюзы, и вся вода, накопленная плотиной, освобождается одновременно. Периодическая смена дня и ночи делает закономерным и естественными описанное выше.
Оптимальными для возникновения жизни являются планеты с продолжительностью планетарных суток в интервале значений 18-48 земных часов. При меньшей продолжительности планетарных суток, описанные выше процессы не достигают уровня, при котором происходит активное движение атмосферных масс и разряды атмосферного электричества, без чего, возникновение органической жизни невозможно. Более длительные планетарные сутки (больше, чем 48 земных часов) приводят к постоянному штормовому состоянию атмосферы планеты, что создаёт тяжёлые условия для возникновения и развития жизни. На таких планетах жизнь может возникнуть только, когда интенсивность излучений звезды, достигающих поверхности планеты, уменьшится до определённого уровня. Только при уровне излучений звезды, когда освещённая поверхность планеты не перегревается, возникают условия для зарождения жизни. Обычно такие условия появляются на последней стадии эволюции звёзд и даже, если на них и возникает жизнь, то она не успевает развиться до сложных форм перед тем, как звезда погибает. Кроме этого, если продолжительность планетарных суток небольшая, перепад мерности не достигает уровня, при котором возникают какие-либо существенные движения масс нижних слоёв атмосферы планеты. Если же продолжительность планетарных суток большая, перепад мерности становится настолько существенным, что приводит к мощным и продолжительным атмосферным бурям и штормам, в результате которых, уничтожается верхний слой планетарного грунта, что создаёт невозможность развития флоры планеты, без которой развитие экологической системы просто невозможно. Штормовое состояние атмосферы вызывает также мощное движение поверхностных слоёв океанов планеты, что, в свою очередь, делает невозможным зарождение жизни в воде.
5. Наличие разрядов атмосферного электричества. Во время разрядов атмосферного электричества, в мрской воде происходит синтез органических молекул. В зоне разряда создаётся дополнительное искривление пространства (изменение уровня мерности), при котором молекулы неорганических соединений, растворённых в воде, соединяются между собой в качественно новом порядке, образуя органические соединения, которые представляют собой цепочки однотипных атомов. Только мощные разряды атмосферного электричества способны создать необходимые условия, при которых уровень мерности достигает критической величины. Две свободные электронные связи каждого из этих атомов в состоянии присоединить к себе, как свободные ионы, так и другие цепочки-молекулы. Атмосферные электрические разряды возникают, как следствие перепада толщины качественного барьера между физическим и вторым уровнями планеты. Когда ночь своим покровом обнимает землю, поверхностный слой планеты начинает охлаждаться и излучать тепловые волны. И, как при всяком излучении, уровень мерности излучающего атома или молекулы уменьшается. Когда это происходит одновременно с триллионами триллионов атомов и молекул на ограниченной территории (площадь, освещённая звездой в дневное время), уровень мерности уменьшается на всей этой территории. Если за день атмосфера и поверхность планеты сильно разогрелись, а ночью произошло резкое охлаждение, возникает скачок уровня мерности. При этом, скопившиеся на уровне качественного барьера свободные материи лавиной обрушиваются вниз. Происходит электрический разряд между атмосферой и поверхностью планеты.
Итак, необходимыми условиями для возникновения жизни на планетах являются:
наличие постоянного перепада мерности,
наличие воды,
наличие атмосферы,
наличие периодической смены дня и ночи,
наличие разрядов атмосферного электричества.
Жизнь зарождается автоматически на всех планетах, где существуют перечисленные выше условия. И таких планет во Вселенной — миллиарды. Наша планета Земля не является уникальным творением природы.
4.3. Качественные особенности органических
молекул и их роль при зарождении жизни
А сейчас рассмотрим, как при перечисленных выше необходимых условиях, зарождается и развивается жизнь. Морская вода, как всем известно, стала колыбелью жизни. В ней содержатся практически все химические элементы и многие соединения из них. Во время разрядов атмосферного электричества происходит деформация пространства. В воде, пронизываемой этими разрядами (молниями), возникает уровень мерности, при котором четырёхвалентные элементы (углерод, кремний, фосфор) начинают соединяться в цепочки. При этом, возникшие молекулы имеют не только структурные отличия, но приобретают и новые качества. Какие же новые качества возникают, при соединении тех же самых атомов в другом структурном порядке? Что заставляет нас разделять атомы, образующие один структурный порядок, от тех же самых атомов, создающих другой структурный порядок? Почему, в одном случае — неорганические соединения, а в другом — органические?
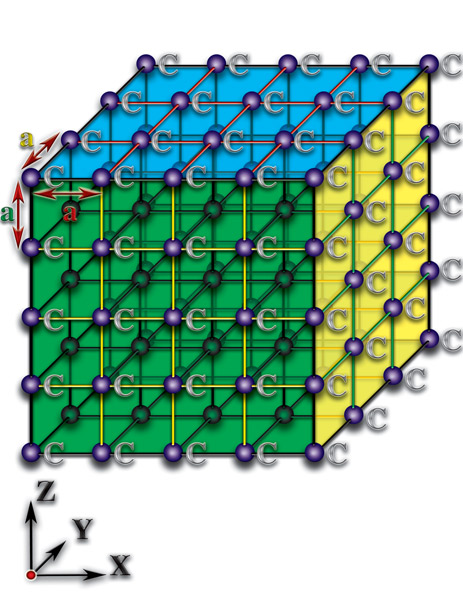
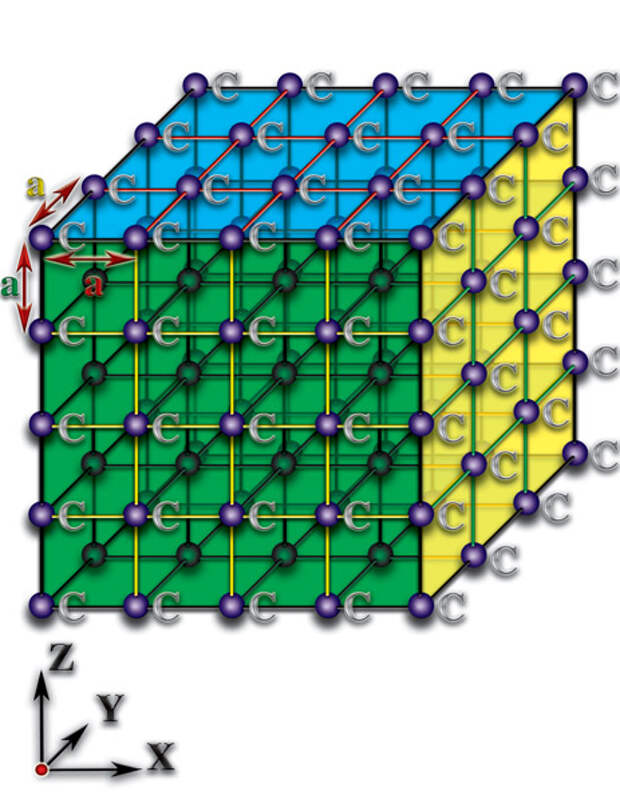
В силу того, что основой белковой жизни является углерод, достаточно проанализировать качественное отличие пространственных характеристик молекул, которые создаёт данный элемент, чтобы разгадать тайну зарождения жизни. Давайте попытаемся понять, к чему приводят различия структурной организации молекул. Рассмотрим неорганические структурные образования — кристаллы. Кристаллы представляют собой такие пространственные соединения, где атомы расположены друг относительно друга на практически одинаковых расстояниях. Эти расстояния соизмеримы с размерами самих атомов (10-14…10-12 метра). Причём, они (расстояния) практически одинаковы по всем пространственным направлениям (алмаз) или тождественны в каждой из пространственных плоскостей (графит). Эти кристаллы образованы атомами углерода (С), но они не являются основой не только живых организмов, но и органических молекул (Рис. 4.3.1
{kind=link}
{kind=link}
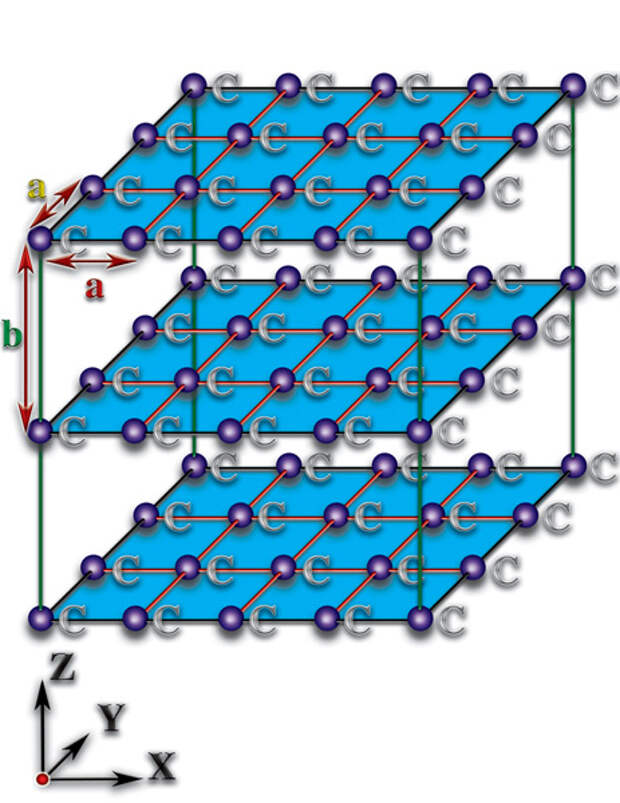
). В чём причины того, что такие же атомы углерода, соединившись в другом пространственном порядке, стали фундаментом живой природы? А они (причины) — следствия качественных особенностей органических молекул (Рис. 4.3.3
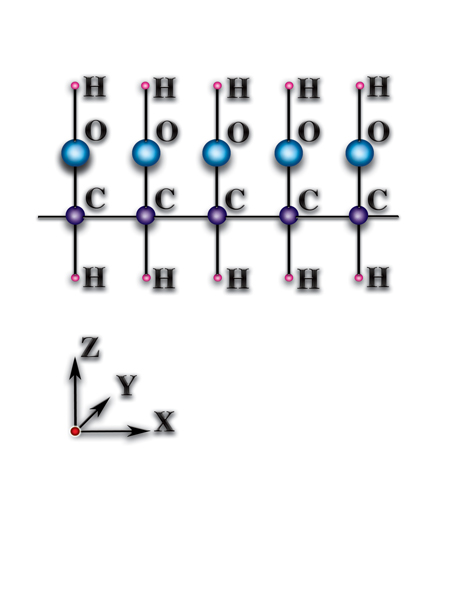{kind=link}
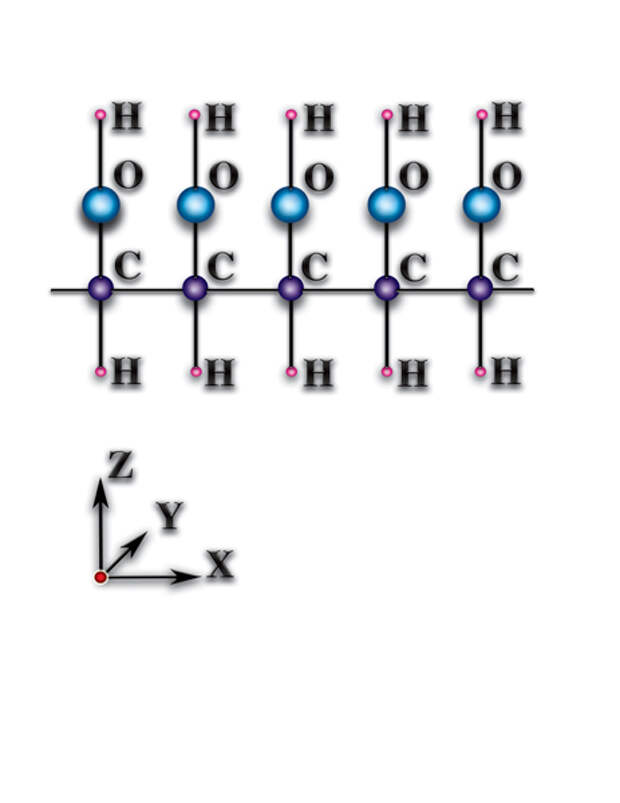
{kind=link}
). Качественные особенности органических молекул следующие:
1. Пространственная структура органических молекул неоднородна в разных пространственных направлениях.
2. Молекулярный вес органических молекул колеблется от нескольких десятков до нескольких миллионов атомных единиц.
3. Неравномерность распределения молекулярного веса органических молекул по разным пространственным направлениям.
И, как следствие перечисленных качественных особенностей, органические молекулы влияют неодинаково на окружающее их микропространство в разных пространственных направлениях. Особенно ярко это явление выражено у молекул РНК и ДНК (Рис. 4.3.5
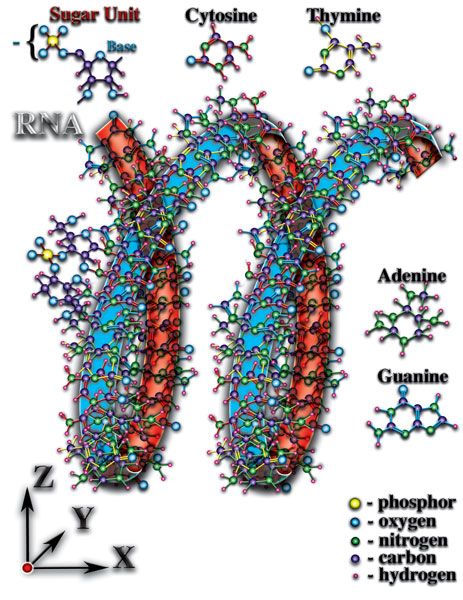{kind=link}
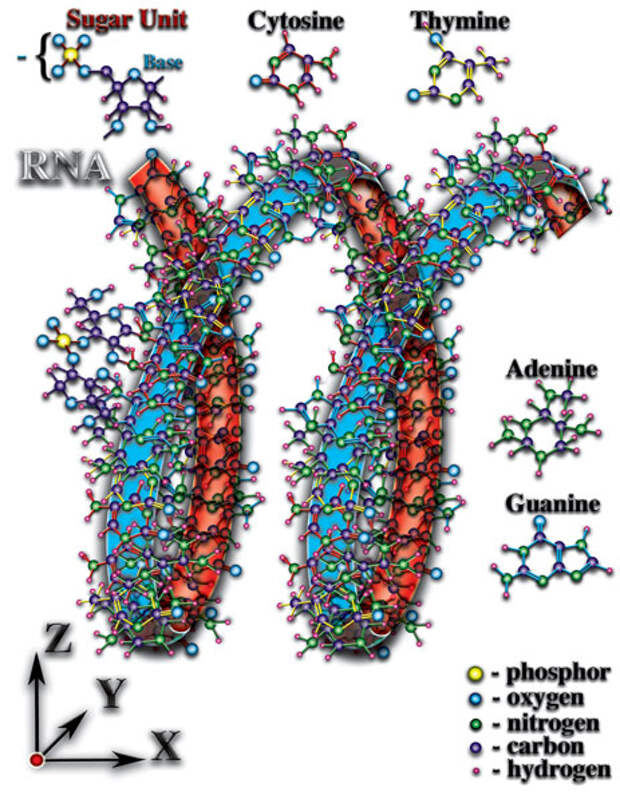
{kind=link}
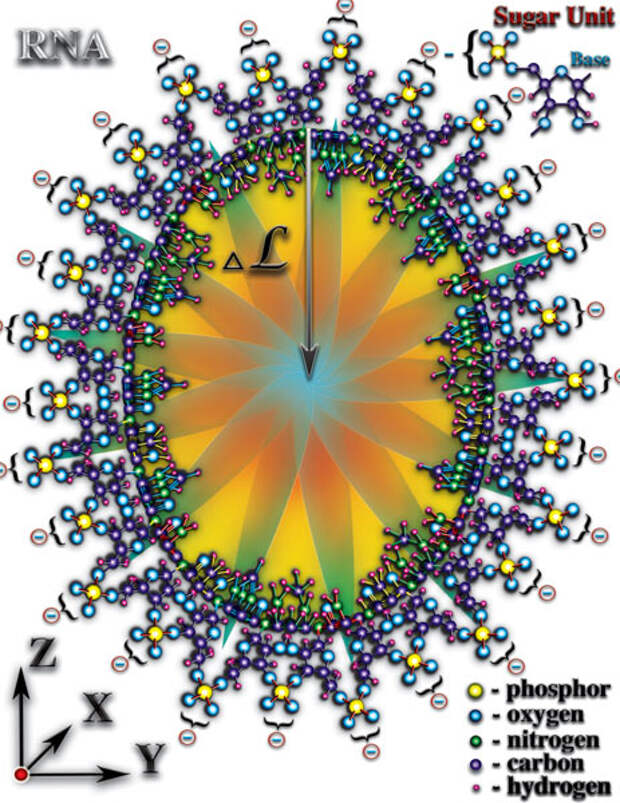
). Атомы, образующие эти молекулы, создают длинные цепочки, закрученные в спираль. Именно спиральная пространственная форма молекул РНК и ДНК создаёт необходимые качества для возникновения живой материи. Какие же это необходимые качества созидают чудо жизни? Что позволяет говорить о качественно новом этапе эволюции материи — эволюции живой материи, эволюции жизни? Попытаемся понять чудо, которое рождает жизнь...
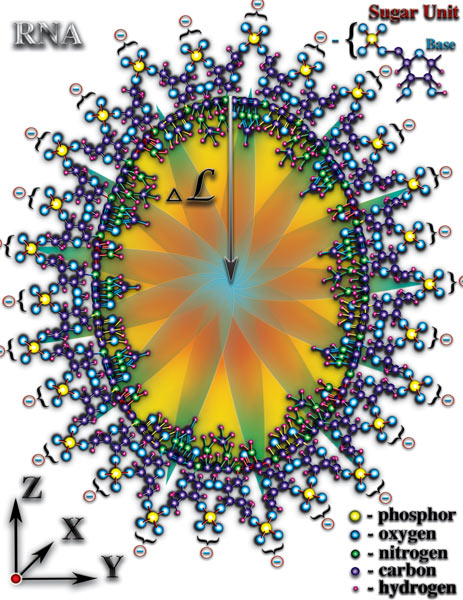
Внутренний объём спиралей молекул РНК или ДНК образует своеобразный туннель. Спиральная молекула оказывает сильное влияние на уровень мерности микропространства этого туннеля. Причём, это влияние на внутренний объём туннеля не одинаково в разных пространственных направлениях (Рис. 4.3.7
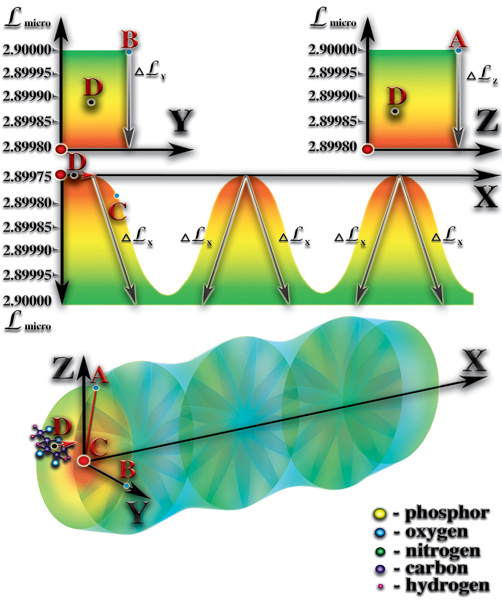{kind=link}
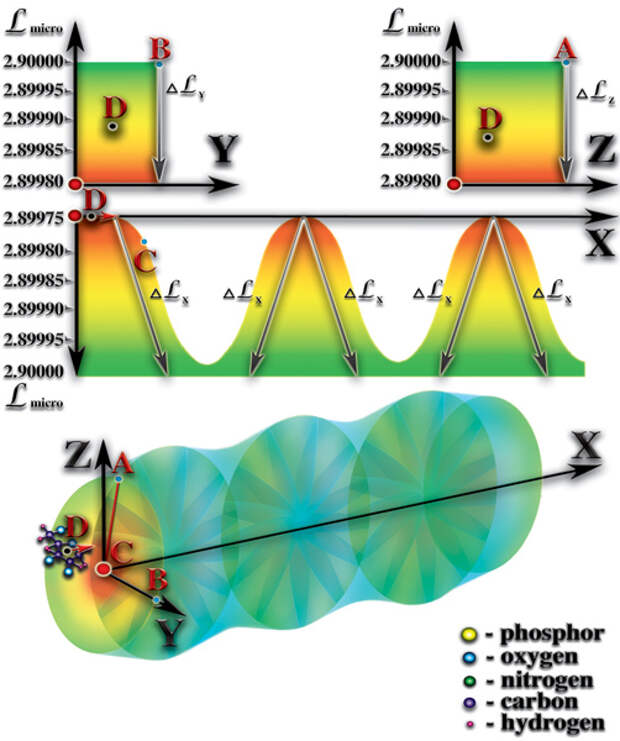
). Вспомним, что каждый атом оказывает влияние на мерность микропространства вокруг себя. Соединение из атомов создаёт комбинацию влияний всех атомов, образующих это соединение, на мерность микропространства молекулы. При этом, важное значение имеет пространственная ориентация влияния каждого атома, входящего в соединение. Спиральная структура молекул РНК или ДНК создаёт условия, при которых влияния на мерность большинства образующих их атомов сосредотачивается во внутреннем объёме спиралей этих молекул. Мерность внешнего объёма спиралей молекул РНК или ДНК претерпевает лишь незначительные изменения. Следует отметить, что изменения мерности внутреннего объёма этих спиралей неодинаковы в разных пространственных направлениях. Вдоль оси витки спирали создают периодически повторяющиеся перепады мерности. Эти перепады во внутреннем объёме создают стоячую волну мерности (волна мерности, параметры которой не изменяются во времени и в пространстве). В радиальных направлениях спираль молекулы РНК или ДНК создаёт плавный перепад мерности. Именно стоячая волна мерности, создаваемая спиральной структурой молекулы РНК или ДНК, является достаточным условием возникновения жизни. Постараемся выяснить почему это именно так. Молекулы РНК и ДНК находятся в водной среде. Морская вода, в которой и зародилась первая жизнь, содержит огромное количество молекул, ионов, как неорганического, так и органического происхождения. Все эти молекулы и ионы находятся в постоянном хаотическом движении. В результате этого движения, молекулы и ионы периодически попадают во внутренний объём спирали РНК или ДНК. И рождается чудо жизни!..
Разгадка этого чуда — очень простая. Дело в том, что внутренний объём спирали молекулы РНК или ДНК является ловушкой для всех попавших в него молекул. Радиальный перепад мерности удерживает попавшие в эту ловушку молекулы внутри спирали РНК или ДНК. При этом, радиальный перепад мерности заставляет свободные материи двигаться вдоль этого перепада. И, как следствие, возникают гравитационные силы, направленные к оси спирали РНК или ДНК. Поэтому, все молекулы, попавшие во внутренний объём спирали, в результате броуновского (хаотичного) движения, начинают двигаться вдоль оси спирали. Так же, как и течение реки увлекает за собой всё, что в неё попадает, радиальный перепад увлекает «пленённые» молекулы. Только очень быстрые молекулы могут вырваться из этого плена. При этом, они теряют часть своего потенциала. Все остальные молекулы начинают вынужденно двигаться вдоль оси спирали. Вдоль оси, спираль молекулы РНК или ДНК создаёт, как Вы помните, стоячую волну перепада мерности. При своём вынужденном движении вдоль оси «пленённые» молекулы попадают в зоны с разными мерностями. Каждая из этих молекул имеет собственный уровень мерности, при котором она максимально стабильна, а также диапазон значений мерности, в пределах которого молекула может существовать не распадаясь. И, как только «пленённые» молекулы, при своём вынужденном движении вдоль оси, попадают в зону с запредельной для них мерностью, они становятся неустойчивыми и начинают распадаться (Рис. 4.3.8
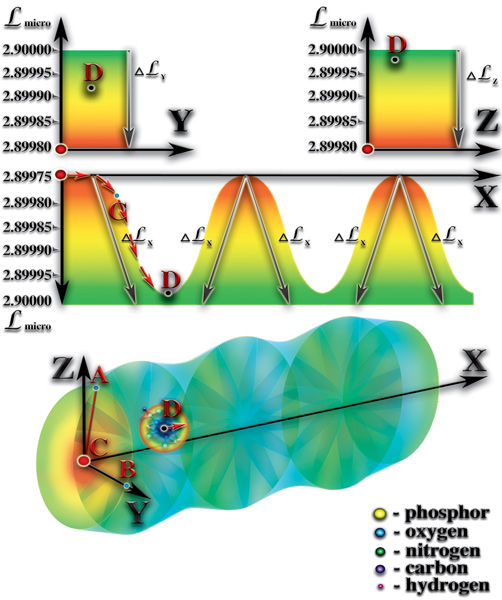{kind=link}

).
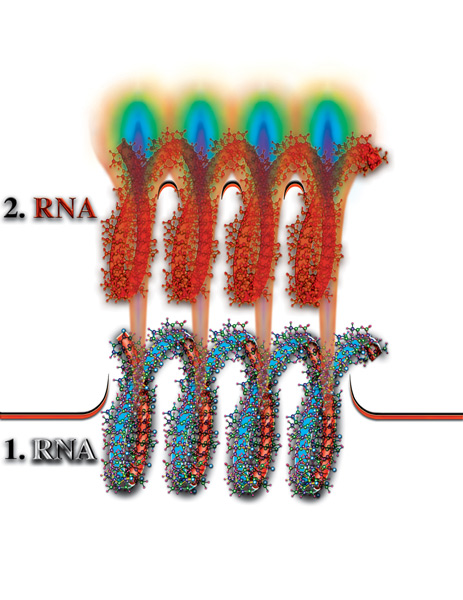
В результате распада молекул, высвобождаются все семь первичных материй, которые образовали физически плотное вещество. При этом, часть высвободившихся материй вновь создаёт новые атомы и молекулы, имеющие собственный уровень мерности, тождественный мерности зоны распада. Обычно вновь возникшие молекулы, при своём вынужденном движении вдоль оси, не распадаются. Выйдя из внутреннего объёма спирали молекулы РНК или ДНК, они оказываются в водной среде (Рис. 4.3.9
{kind=link}
). Эти молекулы часто химически активны и, как следствие, агрессивны, как по отношению к молекулам РНК или ДНК, так и к другим внутриклеточным образованиям. Забегая вперёд, отметим, что эти молекулы, которые в дальнейшем будем называть токсинами или шлаками, выводятся за пределы клетки и далее за пределы организма (в случае многоклеточного организма). Вернёмся к анализу процессов, происходящих во внутреннем объёме спирали РНК или ДНК... Часть высвободившихся свободных материй, как выяснилось, образуют устойчивые атомы и молекулы. А другая часть? Что происходит с ней?!
Именно в этой точке анализа мы подошли к пониманию тайны жизни.
Несвязанные материи, через канал между физическим (первая материальная сфера) и вторым (вторая материальная сфера) уровнями планеты, который возникает во внутреннем объёме спирали РНК или ДНК, начинают перетекать на другие уровни. Вспомним, что каждая молекула, особенно такие огромные, как РНК и ДНК, деформируют микропространство вокруг себя. И при этом деформируется второй материальный уровень планеты. Причём, форма деформации полностью копирует форму молекулы РНК или ДНК, как впрочем и всех остальных молекул. Когда на дороге возникают ямы (деформации), во время дождя они заполняются до краёв водой. Если дождь — продолжительный, дождевая вода, заполнив ямы, начинает стекать в низины. Также и несвязанные материи, перетекая по каналу на второй материальный уровень, полностью заполняют форму деформации. Избыток их возвращает себе свободу от плена планеты. Возникает только один вопрос — какие высвободившиеся материи и почему заполняют эту форму деформации второго материального уровня (сферы)?
Для того, чтобы ответить на этот вопрос, вспомним, что второй материальный уровень (сфера) образовался в результате слияния шести свободных форм материи. Поэтому, деформацию второго материального уровня заполняет только материя G, которая является седьмой материей и не входит в состав гибридной материи второй материальной сферы. После полного заполнения деформации материей G, на втором материальном уровне (сфере) образуется точная копия молекулы РНК или ДНК. Возникает, так называемое, второе материальное тело молекулы РНК или ДНК (Рис. 4.3.10
{kind=link}
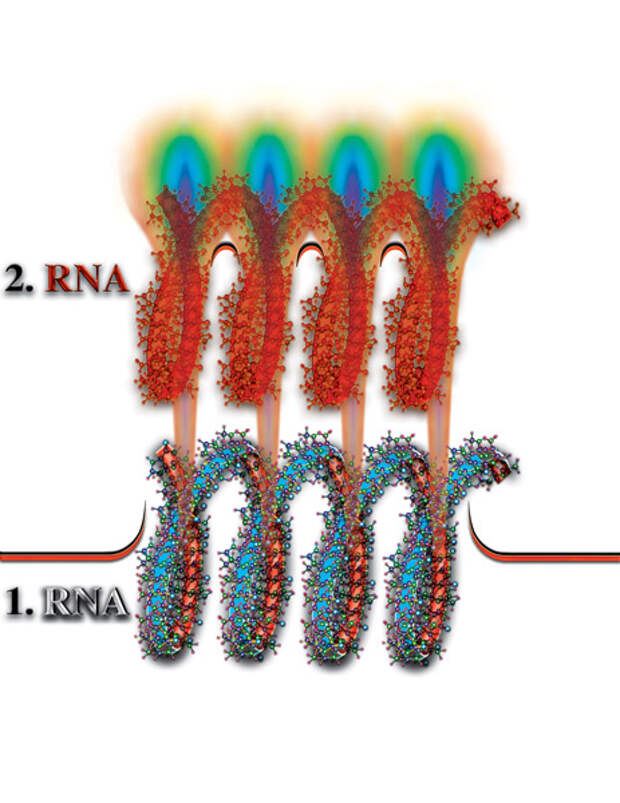
). При полноценном втором материальном теле, между ним — второй материальной сферой и физически плотном телом (первая материальная сфера) исчезает качественный барьер, так как система второе материальное тело, плюс вторая материальная сфера структурно и качественно соответствует физически плотной материи («The DNA PHANTOM EFFECT: Direct Measurement of A New Field in the Vacuum Substructure», by Dr. Vladimir Poponin, 1996.). Между физически плотной (первое материальное тело) молекулой и вторым материальным телом молекулы РНК или ДНК, образуется постоянный канал, по которому высвободившиеся материи продолжают перетекать на второй и другие материальные уровни планеты. Если процесс распада «пленённых» молекул во внутреннем объёме спирали молекулы РНК или ДНК прекратится, то второе материальное тело молекулы или исчезнет совсем, или утратит оптимальную плотность. Как и лужа на дороге: если не будет нового дождя, вся вода из неё испарится, и на дороге останется только яма...
Таким образом, постоянный распад «пленённых» молекул во внутреннем объёме спирали молекулы РНК или ДНК является необходимым условием для поддержания жизни. Возникновение второго материального тела является качественно новой ступенькой в эволюции материи. Пленённая материя нашла способ своего освобождения из своей тюрьмы. И это освобождение — живая материя.
Появление второго материального тела — начало эволюции живой материи.
Первыми живыми организмами стали вирусы. Вирус представляет собой молекулу РНК, окружённую белковой оболочной. Белковая оболочка создаёт вокруг молекулы РНК устойчивую среду, создаётся своеобразный микроклимат вокруг молекулы РНК вследствие того, что белковая оболочка замедляет движение молекул, как внутрь себя, так и из себя. Поэтому, молекулы попавшие внутрь белковой оболочки, сталкиваясь с ней при своём движении, могут много раз срикошетить от неё, перед тем, как покинуть внутренний объём белковой оболочки. Повторные многократные движения попавших внутрь белковой оболочки молекул увеличивают вероятность того, что они попадут в «сферу влияния» молекулы РНК и, как следствие, будут втянуты во внутренний объём молекулы РНК и начнут своё вынужденное движение вдоль оптической оси этой молекулы, попадая под действие стоячей волны мерности. Что, в конечном итоге, и приводит к их распаду на материи, их образующие. Внутренний объём молекулы РНК, как пылесос, засасывает в себя все молекулы, попавшие под воздействие радиального перепада мерности, создаваемого спиралью молекулы РНК. Так же, как и молекулы РНК, «чёрные дыры» макрокосмоса создают вокруг себя сферу влияния, попадая в пределы которой любая материя, в том числе и электромагнитные волны, не в состоянии вырваться. Чёрная дыра макрокосмоса создаёт вокруг себя мощное радиальное гравитационное поле (радиальный перепад мерности), вызывающее распад любой материи. Аналогично внутренний объём спирали молекулы РНК или ДНК, создаёт подобные условия, приводящие к распаду пленённых молекул под действием стоячей волны мерности. Спираль этих молекул ведёт себя идентично «чёрной дыре» макрокосмоса, что позволяет назвать молекулу РНК или ДНК «чёрной дырой» микрокосмоса.
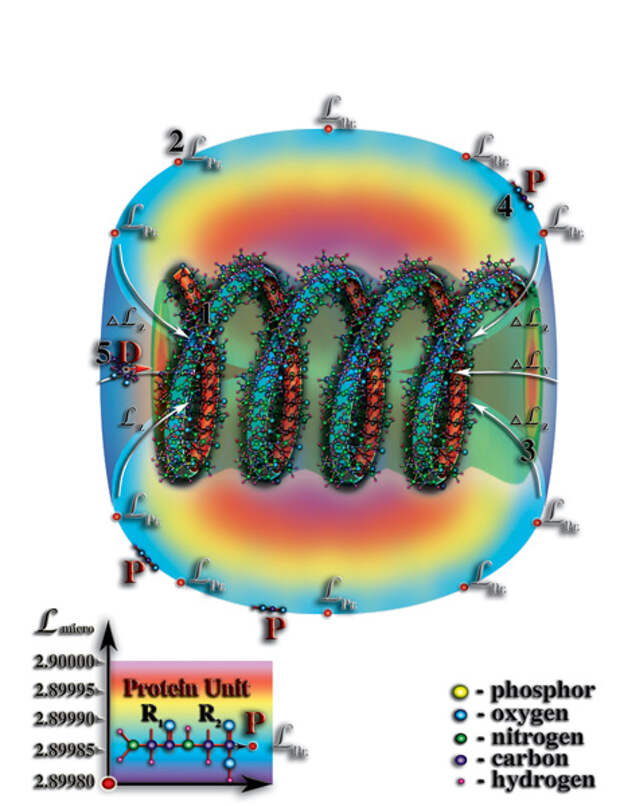
Таким образом, появление белковой оболочки вокруг молекулы РНК явилось следующей ступенью эволюции материи от неживой формы к живой. Именно с появлением этой оболочки, можно говорить о качественно новом этапе эволюции материи — этапе эволюции живой материи. Стоячая волна мерности, возникающая во внутреннем объёме молекулы РНК или ДНК, являясь необходимым условием для возникновения жизни, не является достаточным в силу того, что, в первичном океане концентрация органических молекул была очень маленькой. И поэтому, без дополнительного накопления вблизи молекулы РНК органических молекул не могло быть и речи о постоянном захвате их этой молекулой с последующим распадом на материи, их образующие. Возникает вопрос, каким образом у молекулы РНК могла появиться белковая оболочка? Какое чудо должно было произойти, чтобы это случилось? Ответ на эти вопросы, как ни странно, очень простой. Белки, как все органические молекулы, возникли в насыщенном растворе первичного океана, как следствие атмосферных электрических разрядов. Сами по себе белки являются большими молекулами, состоящиими порой из десятков тысяч атомов и при своём свободном (броуновском) движении в водах первичного океана, попадая в «поле тяготения» молекулы РНК, не могут быть втянуты во внутренний объём этой молекулы в силу того, что белки, сами по себе, представляют огромные молекулы и просто не могут поместиться во внутреннем объёме молекулы РНК. Но это не означает, что молекула РНК не влияет на молекулы белков. Создаваемый спиралью молекулы РНК радиальный перепад мерности, тем не менее, захватывает своим «тяготением» первичные структуры молекул белков и делает их своими постоянными «спутниками», как это происходит при захвате большим материальным телом (например, планетой) меньшего (Рис. 4.3.11
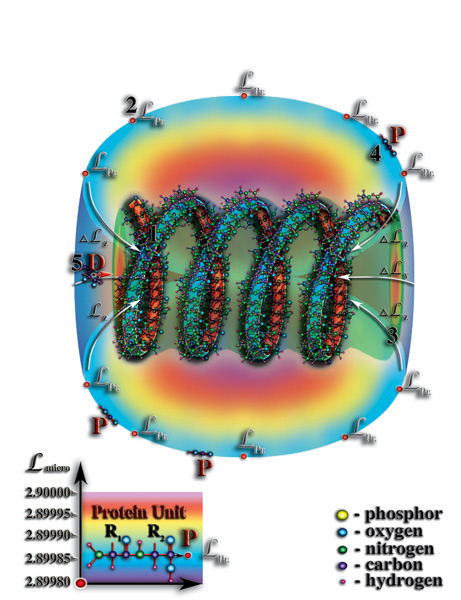{kind=link}
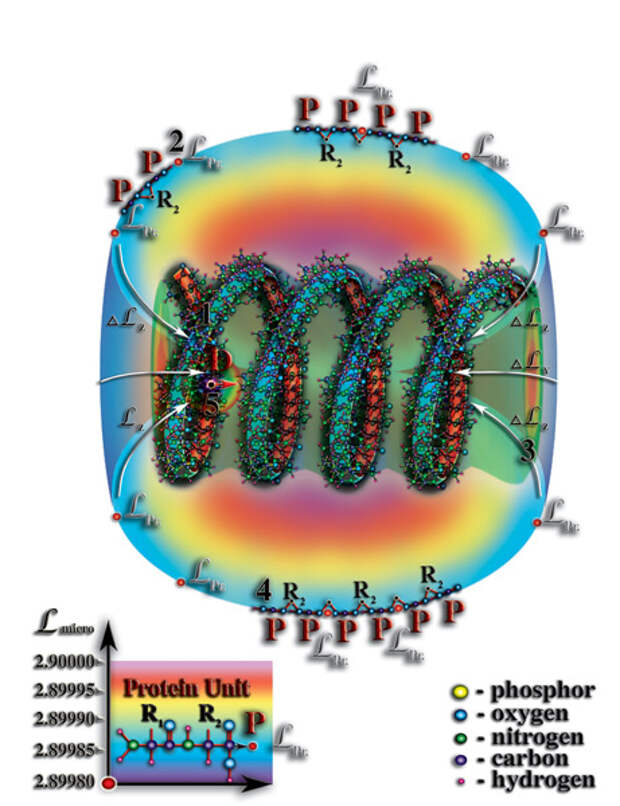
). И опять наблюдается удивительная параллель между макромиром и микромиром. Итак, захват и удержание молекулой РНК первичных структур молекул белков и стало тем условием, приведшим к формированию вокруг этой молекулы белковой оболочки. Со временем, число белков-спутников молекулы РНК возрастало, их «орбиты» были соизмеримы, в силу близких значений уровней собственной мерности. В результате чего, соседние белки-спутники оказывались друг от друга на таком расстоянии, что возникали условия для возникновения, так называемых, химических реакций между ними (Рис. 4.3.12
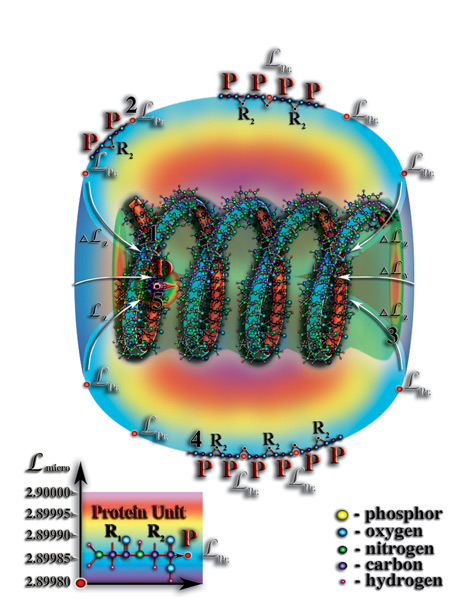{kind=link}
).
Химические реакции между первичными структурами белков-спутников приводили к появлению устойчивых электронных связей между ними и они срастались в одно целое. Постепенно, вокруг молекулы РНК возникла сплошная белковая оболочка. Белки-спутники, в результате этого, заключили в свою собственную «тюрьму» своего захватчика — молекулу РНК (Рис. 4.3.13
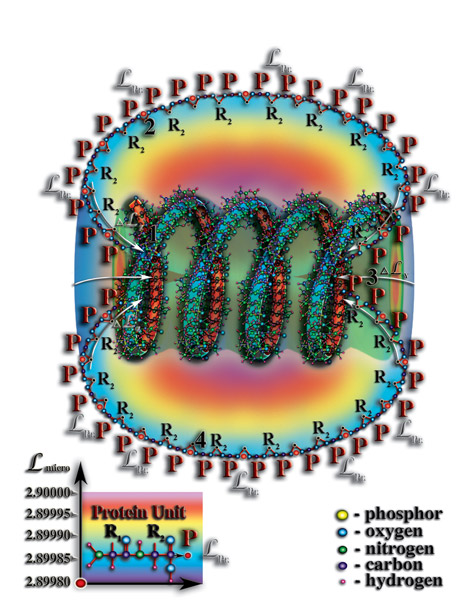{kind=link}

). Таким образом, возникла устойчивая система молекул — молекула РНК и её белковая оболочка. Возник первый живой организм — вирус. Теперь, пришла пора рассмотреть ещё одно качество живой материи — размножение. На уровне вирусов можно говорить о дублировании ими самих себя, в результате чего появление одного живого организма стало естественным результатом жизнедеятельности другого.
Давайте более подробно разберёмся с природой этого явления. Возникшая вокруг молекулы РНК единая белковая оболочка, не является сплошной, по своей сути, а предствавляет собой сетку вокруг молекулы РНК. Ячейки этой «сетки» — не одинаковы, что позволяет молекулам разных размеров попадать внутрь белковой оболочки. Небольшие по размерам молекулы, большинство которых неорганические, довольно свободно могут покинуть внутренний объём белковой оболочки, при своём хаотичном движении, так как большинство ячеек белковой оболочки вируса превышают их размеры. В то время, как большие и средние органические молекулы задерживаются этим «неводом» внутри белковой оболочки, так как вероятность того, что данная молекула попадёт, при своём хаотичном движении в ту же самую ячейку, через которую она попала внутрь, очень мала. В результате этого, внутри белковой оболочки вируса происходит накопление органических молекул. Происходит своеобразная фильтрация воды первичного океана через внутренний объём белковой оболочки вируса. Данный процесс можно рассматривать, как первичное питание первого живого организма. Часть пленённых, подобным образом, органических молекул, попадает в пределы действия радиального перепада мерности спирали молекулы РНК вируса, втягивается во внутренний объём спирали и распадается там на материи, их образующие. Если вода первичного океана в достаточной степени насыщена органическими молекулами, постепенно происходит увеличение концентрации органических молекул внутри белковой оболочки. При росте концентрации органических молекул внутри белковой оболочки, увеличивается и число молекул, которые попадают в «зону притяжения» «чёрной дыры» микрокосмоса — внутренний объём спирали молекулы РНК.
Следует отметить, что внутри белковой оболочки накапливаются разнообразные органические молекулы, в том числе и нуклеотиды — строительный материал для молекул РНК и ДНК. Постепенно концентрация органических молекул внутри белковой оболочки достигает такого уровня плотности, при котором происходит постоянный распад попавших во внутренний объём органических молекул. Вследствие чего, возрастает поток высвобождающихся при распаде первичных материй с физически плотного уровня на второй материальный уровень. Что приводит к избыточному насыщению второго материального тела молекулы РНК. Избыточное насыщение второго материального тела, приводит к появлению обратного потока первичной материи G со второго материального уровня на первый. В результате этого на физическом уровне появляется проекция второго материального тела. Среди органических молекул, насыщающих внутренний объём белковой оболочки вируса, присутствуют и нуклеотиды, которые являются строительным материалом для молекул РНК и ДНК. Поэтому, при появлении проекции второго материального тела, на физическом уровне возникают необходимые условия для соединения отдельных нуклеотидов в спираль молекулы РНК вируса.
Проекция второго материального тела на физическом уровне соответствует точному порядку следования нуклеотидов в молекуле РНК, поэтому она создаёт на физическом уровне дополнительные искривления мерности микропространства, в соответствии с качественными характеристиками соответствующих нуклеотидов, образующих изначальную, так называемую, материнскую молекулу РНК. Рядом с материнской молекулой РНК возникает точная её матрица. Свободные нуклеотиды, попадая в эту матрицу, начинают соединяться друг с другом в том же самом порядке, как и у материнской молекулы РНК. Наведённая матрица вынуждает свободные нуклеотиды соединяться в заданном порядке. Так как, проекция второго материального тела на физическом уровне создаёт такое изменение мерности микропространства, при котором свободные молекулы нуклеотидов не могут соединиться в другом порядке. В результате этого вынужденного соединения свободных молекул нуклеотидов, на физическом уровне появляется новая молекула РНК, которая является точной копией материнской.
Но почему нуклеотиды не могут соединиться в другом порядке? Ответ на этот вопрос — очень простой. Каждый нуклеотид имеет собственный уровень мерности, отличный от другого, поэтому, для того, чтобы два нуклеотида соединились друг с другом, необходимо создать дополнительное изменение мерности микропространства. Причём, для разных пар нуклеотидов, величина этого дополнительного изменения мерности микропространства будет другой. Поэтому, когда на физическом уровне возникает достаточно плотная проекция второго материального тела молекулы РНК вируса, изначальный уровень мерности микропространства в зоне проекции изменяется в точном соответствии с кодом данной молекулы РНК данного вида. Что и приводит к тому, что только тот нуклеотид, параметры которого тождественны данной области микропространства только и может «занять» данное место. Каждый нуклеотид буквально «садится» в специально подготовленное для него «гнездо». В результате этого процесса, появляются две тождественные молекулы РНК данного вируса. А после этого наступает этап создания новой молекулой РНК белковой оболочки, по тому же принципу, что у материнской молекулы РНК вируса из белков, накопленных в белковой оболочке материнского вируса. Подобный процесс будет возникать каждый раз, когда внутри белковой оболочки вируса накопится достаточное количество органических молекул — «строительный материал» нужного качества. Происходит процесс дублирования (по-другому — размножение) вируса. В результате этого процесса, органическая материя, возникшая в первичном океане из неорганической, под воздействием атмосферных электрических разрядов реорганизуется в простейшую самоорганизующуюся живую материю. Таким образом, появились первые примитивнейшие живые организмы — вирусы, эволюция которых и привела к многообразию растительных и животных форм жизни сначала в мировом океане, а потом и на суше.
Следующей эволюционной ступенькой развития жизни стали, так называемые, бактериофаги, промежуточное звено между вирусами и бактериями — простейшими одноклеточными организмами. Может возникнуть вопрос: каким образом произошла дальнейшая эволюция жизни от вируса — к бактериофагу, от бактериофага — к одноклеточному организму? И опять нет места никакому чуду, всё — очень просто и в тоже время прекрасно. Размножившиеся вирусы заполнили верхний слой первобытного океана на глубину не более ста метров. Данная глубина (сто метров) проникновения вирусов в мировой океан обусловлена тем, что синтез органических молекул происходит во время атмосферных разрядов электричества, которые затрагивали только поверхностный слой первичного океана. Это, во-первых, а, во-вторых, именно верхний слой океана находился в постоянном движении, под воздействием ветров и приливов, и на эту глубину проникает солнечный свет. Так вот, вирусы «плавающие» в поверхностном слое первичного океана периодически оказывались в зоне действия атмосферных разрядов электричества. Атмосферные разряды вызывают изменение мерности пространства в зонах своего прохождения, создавая, тем амым, условия для синтеза органических соединений. Но, что произойдёт, если в зоне электрического разряда окажется вирус? Конечно, если вирус попадёт под прямое воздействие электрического разряда, произойдёт полное его разрушение. Что же произойдёт если вирус попадёт в периферийные зоны воздействия атмосферного разряда? Произойдут ли, при этом, какие-либо изменения? При изменении мерности пространства вокруг и внутри вируса, возможно протекание нескольких процессов:
1. Изменение порядка соединения нуклеотидов в существующей молекуле РНК вируса.
2. Увеличение или уменьшение числа нуклеотидов в существующей молекуле РНК вируса.
3. Появление химических связей между существующей молекулой РНК вируса и другими молекулами РНК, которые находились в момент электрического разряда внутри белковой оболочки вируса или появились в ней в результате воздействия электрического разряда.
При изменении порядка соединения нуклеотидов в молекуле РНК вируса, появляется новый вирус, как результат, так называемых, мутаций. При уменьшении числа нуклеотидов, образующих молекулу РНК вируса, последний может утратить качества живой материи, так как, для проявления свойств живой материи, молекула РНК должна достичь критического молекулярного веса. Вирусы являются пограничной формой организации материи между живой и неживой. Для проявления свойств живой материи, молекула РНК вируса должна достичь определённой молекулярной массы, при которой возникает эффект открытия качественного барьера между первой и второй материальными сферами. При меньшей молекулярной массе, качественный барьер не открывается. Именно поэтому, если вирус удалить из воды, он переходит в кристаллическое состояние, так как вне воды молекула РНК вируса теряет группы H и OH со своих внешних электронных связей, что довольно сильно сказывается на её молекулярном весе и, как следствие, качественный барьер восстанавливается и исчезают свойства живой материи. Попадая в воду, молекула РНК вируса восстанавливает свои химические связи, в результате чего, группы H и OH присоединяются к ней, и молекулярный вес вновь возрастает. При достижении критического молекулярного веса происходит открытие качественного барьера между первой и второй материальными сферами, и вновь появляются свойства и качества живой материи. Таким образом, весьма важным фактором, определяющим возможность зарождения жизни, является молекулярный вес, точнее, существует качественная граница молекулярного веса молекул РНК или ДНК, так называемый, критический молекулярный вес, при котором эволюция материи выходит на качественно новую ступень эволюции — эволюцию живой материи.
Если же под воздействием атмосферных разрядов электричества увеличивается число нуклеотидов в молекуле РНК вируса, наблюдается несколько любопытных моментов. Во-первых, появление «лишних» нуклеотидов приводит к рождению нового вируса, новой мутации. Во-вторых, увеличение числа нуклеотидов приводит к росту молекулярного веса молекулы РНК, вследствие чего увеличивается степень её влияния на окружающее микропространство, что приводит в свою очередь к увеличению размера белковой оболочки. Увеличение размеров белковой оболочки связано с тем, что более тяжёлая молекула РНК вируса в большей степени влияет на окружающее микропространство. Вследствие чего белки-спутники, образующие оболочку вируса, захватываются «полем тяготения» молекулы РНК на большем удалении от неё самой, что приводит к тому, что у более тяжёлой молекулы РНК появляется большего размера белковая оболочка. Большего размера белковая оболочка позволяет накапливать внутри себя больше органических молекул и позволяет создать более устойчивый внутренний микроклимат.Если же, во время атмосферных разрядов электричества, возникают устойчивые химические связи между двумя молекулами РНК, появляется молекула, представляющая собой пространственно-химическое соединение двух спиралей — появляется, так называемая, двуспиральная молекула РНК, при определённых условиях появляется молекула ДНК. Появление ДНК открывает новую эру развития живой материи — от одноклеточных живых организмов до многоклеточных и т.д. до появление разумной живой материи. Двойная спираль молекулы ДНК создаёт более выраженную деформацию микропространства около себя, что ускоряет процесс распада «пленённых» молекул на материи их образующие, в силу того, что при своём вынужденном движении во внутреннем объёме спиралей молекулы ДНК «пленённые» молекулы попадают под удары двух стоячих волн мерности, в то время, как у молекул РНК имеется только одна стоячая волна мерности. Двойная стоячая волна мерности молекулы ДНК, таким образом, ускоряет процесс распада «пленённых» молекул, увеличивая тем самым эффективность всей системы в целом. Кроме этого, двойная спираль молекулы ДНК создаёт такое влияние на своё микропространство, что белковая оболочка возникает на значительно большем расстоянии от самой молекулы, что позволяет накопить внутри такой оболочки значительно больше захваченных в «плен» органических молекул.
Это — важнейший фактор для поддержания жизни.
Больших размеров белковая оболочка «процеживает» большее количество морской воды с «плавающими» в ней органическими молекулами, возникающими при атмосферных разрядах электричества. Это естественно: большим неводом можно поймать больше рыбы. Только неводом, в данном случае, является белковая оболочка, а рыбой — органические молекулы, «плавающие» в первичном океане. Кроме этого, двойная спираль молекулы ДНК создаёт условия для появления многослойной оболочки, так называемой, мембраны. Мембрана, в процессе эволюции, сформировалась из трёх слоёв: двух белковых и одного жирового. Причём, жировой слой располагается между двумя белковыми. Спирали молекулы ДНК пространственно смещены одна относительно другой. Поэтому, каждая из этих спиралей создаёт «свои» белковые оболочки, которые оказываются тоже смещёнными друг относительно друга и, кроме этого, одна внутри другой. Формируются две белковые «крепостные» стены вокруг молекулы ДНК. В результате чего, органические и неорганические молекулы вынуждены просачиваться через преграды, чтобы попасть во внутренний объём оболочки. При прохождении через двойную преграду, эти молекулы теряют свою кинетическую энергию. И, как следствие, практически не в состоянии прорваться назад через оболочки. Таким образом, происходит фильтрация воды первичного океана и накопление органических молекул внутри оболочек. Между вложенными, одна в другую, белковыми оболочками образуется зазор. И любая молекула, после прохождения через внешнюю белковую оболочку, попадает в пространство между внешней и внутренней. Собственный уровень мерности белковых оболочек значительно выше собственного уровня воды океана. Поэтому, возникает двойной незначительный перепад мерности с зоной устойчивого равновесия между ними. При своём движении, все молекулы должны преодолеть эти перепады мерности и попадают в «нейтральную» зону с уровнем мерности меньшим, чем уровень собственной мерности белковых оболочек. Именно поэтому жировые молекулы, попадая в зазор между белковыми оболочками, попадают в зону с уровнем мерности весьма близким к уровню собственной мерности жировых молекул. Жировые молекулы начинают, как бы, оседать в пространстве между белковыми оболочками, постепенно заполняя собой этот зазор. При этом, молекулы жира соединяясь друг с другом, создают жировую прослойку между белковыми оболочками. Со временем, возникают химические связи между жировой прослойкой и белковыми оболочками. И как результат, возникает трёхслойная оболочка — мембрана.
При появлении трёхслойной оболочки, можно говорить о следующей ступени развития живой материи — возникновении одноклеточных организмов. Их преимущество перед вирусами в том, что многослойная клеточная мембрана создаёт внутри клетки устойчивую химическую среду. Кроме этого, клеточная мембрана является защитой от агрессивности внешней среды, что создаёт благоприятные условия для дальнейшей эволюции жизни. Гидрофобные (водоотталкивающие) свойства жирового слоя мембраны создавали благоприятные условия для проникновения внутрь оболочки органических молекул, создавая сложности для проникновения внутрь оболочки молекул воды. Дело в том, что внутренний объём оболочки — ограничен и поэтому, если в него попадает молекула воды, которая занимает пусть небольшой, но тем не менее, реальный объём пространства, не остаётся места для органических молекул, которые гораздо больше и, в силу этого, двигаются значительно медленнее молекул воды. Таким образом, появление жирового слоя оболочки практически выравняло шансы органических молекул и неорганических. Подобная оболочка как бы «придерживает» часть молекул воды, создавая благоприятные условия для проникновения органических молекул. Это — огромнейшее приобретение, но, как и за всякое приобретение, за него одноклеточные организмы были вынуждены платить большую цену. Если вирусы могут существовать миллионы и миллиарды лет, периодически находясь в живом или кристаллическом состоянии, то одноклеточные организмы, как потом и многоклеточные, стали «смертными». У «молодого» одноклеточного организма толщина и плотность жирового слоя мембраны относительно невелика, что позволяет молекулам воды попадать внутрь оболочки. Со временем, происходит окисление жировых оболочек, в результате чего гидрофобные свойства мембраны усиливаются, кроме того, со временем жировой слой оболочки как бы «толстеет», как результат продолжения захвата белковыми оболочками новых жировых молекул из окружающей среды. И, как следствие, постепенно замедляется, а потом и полностью прекращается циркуляция веществ через мембрану. При потере внутренним объёмом клетки определённого количества воды, прекращается жизнедеятельность данной клетки, клетка погибает. Таким образом, одноклеточные организмы стали смертными, т.е. могут существовать только ограниченное время.
Появление трёхслойной мембраны дало колоссальный толчок развитию жизни и, в тоже время, возникли временные ограничения продолжительности жизнедеятельности одноклеточных организмов. При потере воды, они, в отличие от вирусов, погибали. Поэтому первые одноклеточные организмы могли существовать только в водах первичного океана.Движение верхних слоёв первичного океана приводило к тому, что однотипные одноклеточные организмы попадали в разные внешние условия. Влияние разных внешних условий на однотипные одноклеточные организмы создавало такие условия, при которых они или погибали, или изменялись. Появились растительные и животные одноклеточные организмы. Многообразие внешних условий порождало многообразие форм растительных и животных организмов. Стала формироваться первичная экологическая система. Способность одноклеточных животных организмов самостоятельно перемещаться, дала новый толчок эволюции жизни. Животные одноклеточные организмы приобрели с этим некоторую независимость от капризов внешней среды. Первобытный океан содержал ещё очень мало органических веществ и первым одноклеточным организмам было весьма сложно «выловить» в окружающей воде органические вещества, которые необходимы для поддержания их жизнедеятельности. Вспомним, при каких условиях из неорганических молекул углерода, кислорода, азота, водорода и других возникают органические соединения... Происходит это, когда насыщенную неорганическими молекулами и атомами воду, пронизывают электрические разряды, возникающие, как результат перепада статического электричества между атмосферой и поверхностью. Электрические разряды искривляют микрокосмос, что и создаёт условия для соединения атомов углерода в цепочки — органические молекулы. Таким образом, чтобы возник синтез органических молекул, необходимо изменение мерности микрокосмоса на некоторую величину:
ΔL ≈ 0,020203236... (4.3.1)
И чтобы первые одноклеточные организмы могли восстанавливать и сохранять свою структуру, необходим синтез простейших органических соединений внутри самих одноклеточных организмов. Возникновение синтеза простейших органических молекул из неорганических возможно при изменении мерности микрокосмоса на величину ΔL/2. Никакой простейший (и даже сложный!) живой организм создать электрический разряд подобный атмосферному не в состоянии. В ходе эволюции у простейших одноклеточных организмов возник промежуточный вариант, дающий необходимую величину ΔL. Вспомним, что каждая молекула, атом влияет, искривляет свой микрокосмос на ту или иную величину. Максимальное влияние на микрокосмос оказывают органические молекулы. Большие органические молекулы, такие как ДНК и РНК оказывают такое влияние на микрокосмос, при котором происходит не синтез, а распад простых органических молекул, под воздействием перепадов мерности, создаваемых стоячей волной мерности внутреннего объёма спирали молекул РНК или ДНК. Вспомним, что синтез органических молекул из неорганических изначально происходил при атмосферных разрядах электричества, которые создавали уровень мерности, необходимый для возможности атомам углерода С соединится в цепочки. Поэтому для возникновения синтеза органических молекул внутри клетки должны происходить процессы аналогичные перечисленным. Клетка не в состоянии создать электрический разряд, аналогичный разрядам атмосферного электричества, но тем не менее, в ней процесс синтеза органических молекул происходит. Каким же образом природа решила эту проблему!? И опять, всё элементарно просто.
Для синтеза органических молекул из неорганических необходимо создать периодические колебания мерности микрокосмоса в пределах 0 < ΔL ≤ 0,020203236, которое будет накладываться на уже существующее искривление пространства, создаваемое клеточными включениями. При этом, происходит накладывание на постоянный уровень мерности периодически меняющейся величины. И на некоторое короткое время в микроскопическом объёме пространства возникают условия так необходимые для синтеза органических молекул. Атмосферные электрические разряды происходят на макроуровне, а синтез органических молекул клетками — на микроуровне. В первом случае синтез является побочным эффектом, во втором — прямым. Для того, чтобы это произошло, необходимо наличие у клетки молекул, собственный уровень мерности которых, плюс периодическое колебание мерности, приходящее извне клетки, создавали бы вместе необходимые условия для синтеза. Такое влияние на микрокосмос оказывают средней величины органические молекулы. Казалось бы, всё очень просто... В одноклеточных организмах должны быть молекулы, примерно, на порядок меньше молекул ДНК и РНК, и проблема уже решена... Но, не всё так просто. Каждая молекула изменяет микрокосмос вокруг себя, но это изменение продолжает быть неизменным до тех пор, пока сохраняется целостность самой молекулы. Для того, чтобы возник синтез органических молекул должно возникнуть колебание мерности микрокосмоса с амплитудой:
0 < ΔL < 0,010101618... (4.3.2)
Колебания мерности микрокосмоса должны быть, по крайней мере периодическими, чтобы возникли нормальные условия для синтеза органических молекул. Для этого, должны быть молекулы, которые бы изменялись при незначительных изменениях внешней среды и вызывали внутри одноклеточных организмов нужные колебания мерности микрокосмоса. Эти воздействия внешней среды (излучения) не должны в то же самое время разрушать сами одноклеточные организмы, но должны свободно попадать внутрь их мембран. Отвечающими всем этим требованиям внешними факторами являются слабые тепловые и оптические излучения Солнца, в то время, как другая часть солнечной радиации для органических соединений и организмов (рентгеновское и гамма-излучения) является разрушающей. И вновь спасение — в воде... Вода океана поглощает рентгеновское и гамма-излучения и пропускает тепловое и оптическое излучения Солнца, которые также свободно могут проникнуть в одноклеточные организмы. Таким образом, для того, чтобы возник внутриклеточный синтез органических соединений, необходимы следующие условия:
а) наличие внутри одноклеточных организмов органических молекул, которые легко изменяют свою структуру в некоторых пределах, при изменении внешних факторов, что приводит к колебанию мерности микрокосмоса в диапазоне 0 < ΔL < 0,010101618...
б) наличие внешних факторов, которые могут вызывать нужные изменения структуры этих молекул, не разрушая молекулы, как и сами одноклеточные организмы (слабые тепловые и оптические излучения Солнца).
В ходе эволюции, возникла нужная для этого молекула — молекула хлорофилла. Молекулы хлорофилла, поглощая часть оптического и теплового излучения, изменяют свою структуру, создавая новые соединения, в свою очередь очень неустойчивые, причём, поглощение происходит порциями, так называемыми, фотонами. Эти соединения распадаются, как только прекращается действие теплового и оптического излучения, и именно это вызывает нужные колебания мерности микрокосмоса, которые так необходимы для возникновения процесса синтеза внутри одноклеточных организмов. Поглощая фотоны солнечного излучения, молекула хлорофилла вызывает колебания мерности микрокосмоса. Это связано с тем, что, при поглощении фотонов атомами молекулы хлорофилла, электроны переходят на другие орбиты. При этом, на возникшие электронные связи, молекула хлорофилла присоединяет группы ОН и Н, что приводит к колебанию молекулярного веса. И, как следствие, колебанию мерности микрокосмоса, что и создаёт необходимые условия для возникновения синтеза органических соединений. Накопленный потенциал молекула хлорофилла теряет во время синтеза и возвращается в исходное, более устойчивое состояние, готовая к новому поглощению фотонов. Синтез происходит с поглощением из окружающей среды углекислого газа (СО2) и, как побочный продукт, выделяется кислород (О2). Происходит, так называемый, фотосинтез. Следовательно, простейшие одноклеточные организмы в ходе эволюционного развития (благодаря молекулам хлорофилла) приобрели способность, поглощая солнечный свет, синтезировать органические соединения, которые необходимы для восстановления их структуры и жизни.
Соединение одноклеточных организмов отростками клеточных мембран в один конгломерат (например, вольвокс) стало причиной очередного эволюционного скачка жизни. Срастание одноклеточных организмов, посредством отростков клеточных мембран, явилось причиной очередного взрыва развития жизни. Временные соединения превратились в постоянный симбиоз одноклеточных организмов. С этого момента эволюции жизни, можно говорить о многоклеточных организмах. Наружные клетки многоклеточного конгломерата подвергались воздействию внешней среды, часто агрессивной, в то время, как внутренние клетки многоклеточного организма своей внешней средой имели окружение из других клеток. В результате этого, со временем клетки многоклеточных организмов стали выполнять разные функции и приобрели разный внешний вид. В ходе эволюции возникали новые виды многоклеточных организмов, исчезали старые. Более совершенные экологические системы приходили на смену простым. Со временем, жизнь выбралась из своей колыбели — океана и освоила сушу. Но всё это происходило на физически плотном уровне. Как же эти эволюционные процессы отражались на других уровнях планеты?..
Вспомним, что молекула РНК или ДНК на втором материальном уровне создаёт свою точную копию из одной материи. Она (копия) является, так называемым, вторым материальным телом этой молекулы. Одноклеточный организм (клетка), кроме молекул ДНК, образующих хромосомы ядра клетки, включает в себя целый ряд органических включений (аппарат Гольджи, митохондрии, центриоли, эндоплазматическая сеть и т.д.), а также, органические и неорганические молекулы. Последние принимают участие во внутриклеточных биохимических реакциях. Так вот, все клеточные включения тоже оказывают влияние (т.е. деформируют, искривляют) на окружающее микропространство. Отличие их влияния от влияния молекул РНК и ДНК состоит в том, что большинство из них (за исключением РНК митохондрий) не открывают качественного барьера между физическим и вторым материальным уровнями. Поэтому на втором материальном уровне все эти деформации, вместе взятые, создают точную копию физически плотной клетки (Рис. 4.3.14
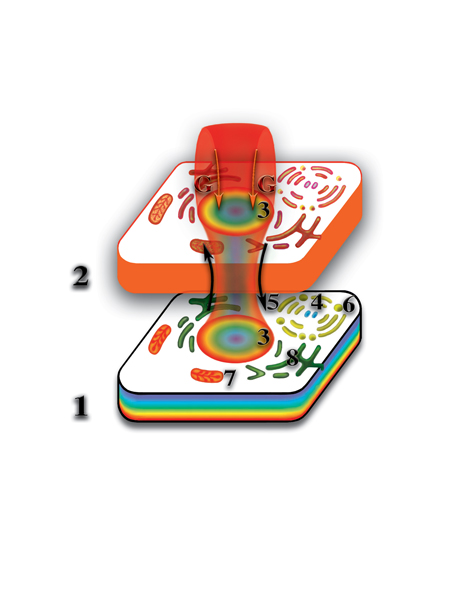{kind=link}
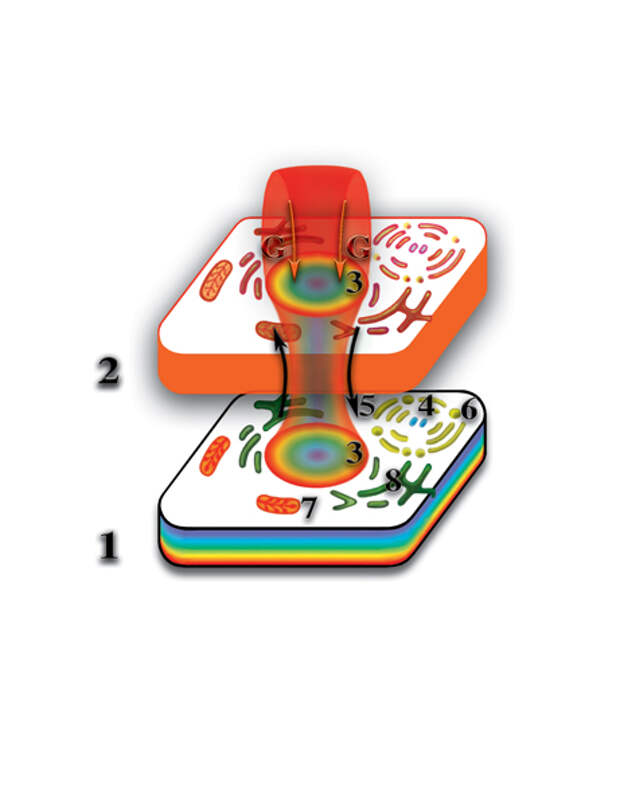
). Так же, как и следы на влажной земле повторяют форму ног, так и второе материальное тело клетки является полной копией физически плотной клетки. Отличие только в том, что второе материальное тело клетки образуется из одной первичной материи, в то время, как физически плотная клетка — слиянием семи первичных материй. Таким образом, образуется система физически плотная клетка — второе материальное тело клетки. В физической клетке постоянно происходят процессы расщепления физически плотного вещества. Первичные материи высвобождаются и начинают циркулировать между уровнями по создаваемому ядром клетки каналу, формируя защитную оболочку клетки.
Как же возникает из выбрасываемых по каналу первичных материй защитная оболочка клетки? Какие природные или божественные силы «позаботились» о такой защите живых созданий?
И вновь, к сожалению многих, никакого божественного начала в этом нет. Всё, как и всегда, и очень просто, и одновременно очень сложно. Хромосомы, образующие ядро клетки, деформируют микропространство вокруг себя. При этом, в зоне деформации увеличивается мерность микропространства. Высвободившиеся при расщеплении первичные материи начинают двигаться по создаваемому ядром клетки каналу с физического уровня на второй, третий материальные уровни и т.д. Этот поток первичных материй направлен против основного потока первичных материй макропространства. Поэтому выбрасываемые через канал клеточного ядра первичные материи разворачиваются во встречных потоках первичных материй, формирующих сферы планеты. Аналогией этому может служит фонтан. Струя воды, выбрасываемая под давлением, поднимается до определённой высоты. Израсходовав начальный потенциал, она спадает вниз, создавая своеобразный водяной купол. Так и первичные материи, выбрасываемые через канал клеточного ядра, разворачиваются встречными потоками. И двигаются вдоль зоны искривления микропространства. Достигнув физического уровня они, повторяя форму искривления микропространства, заворачиваются к клеточному ядру. В результате вокруг физически плотного и второго материального тел клетки первичные материи создают изолированную зону (Рис. 4.3.15
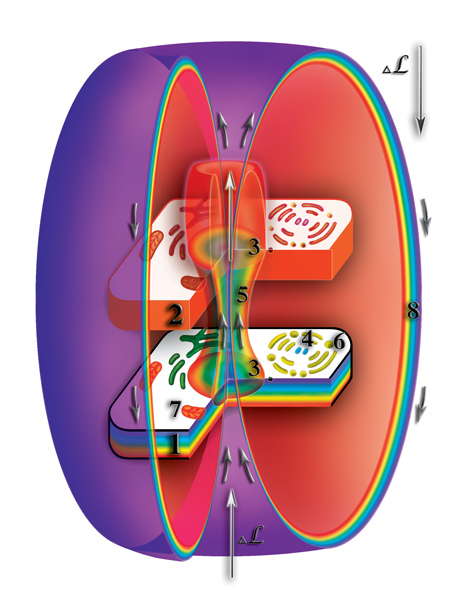{kind=link}
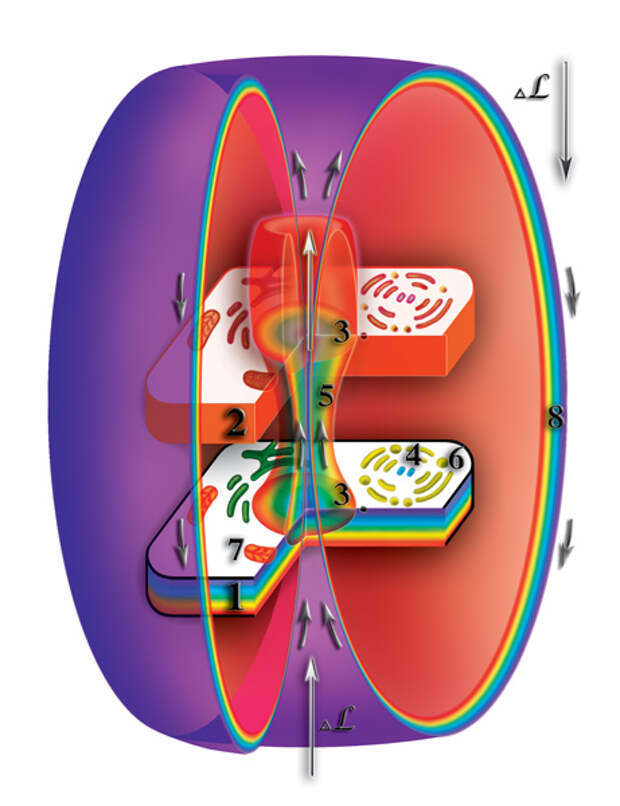
). После завершения формирования защитной оболочки, общий поток первичных материй просто огибает эту зону. Внутри данной защитной оболочки возникает своеобразный микроклимат, оазис, где второе материальное тело клетки максимально изолируется, как от хаоса окружающей среды, так и от влияния других клеток или организмов. Защитная изолирующая оболочка будет существовать до тех пор, пока будет происходить расщепление веществ внутри клетки и функционировать канал между уровнями клетки. Другими словами, до тех пор пока клетка остаётся живой. В многоклеточных организмах клетки имеют различные функции и, как следствие, приобретают разные внешние формы. Любой многоклеточный организм представляет собой жёсткую колонию, в которой внешнюю среду большинства клеток образуют другие клетки того же организма. Причём, это фиксированное положение клеток сохраняется на протяжении всей их жизни (исключение составляют клетки крови).
Вспомним, что каждая живая клетка создаёт второе материальное тело, которое представляет собой её структурную копию. В жёсткой колонии положение клеток зафиксировано, поэтому их вторые материальные тела также имеют фиксированное положение. Поэтому на втором материальном уровне вторые материальные тела клеток образуют аналогичную жёсткую систему — второе материальное тело многоклеточного организма. В ходе эволюции многоклеточных организмов специализация клеток привела не только к тому, что они стали выглядеть по другому, но и степень их влияния на свой микрокосмос претерпела существенные качественные изменения. Деформация микропространства, создаваемая несколькими типами клеток многоклеточного организма привела к тому, что происходит открытие качественного барьера между вторым и третьим материальными уровнями планеты. При этом, на третьем материальном уровне формируются, по аналогии со вторым материальным уровнем, точные копии физически плотных клеток со всеми их особенностями. Назовём эти копии третьими материальными телами физически плотных клеток. Отличие их от вторых материальных тел клеток определяется не только расположением на следующем качественном уровне планеты, но и качественным составом. Полные третьи материальные тела образуются в результате синтеза из двух первичных материй (Рис. 4.3.16
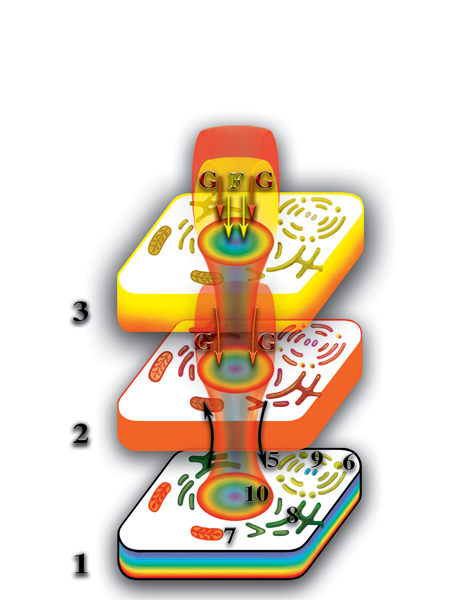{kind=link}
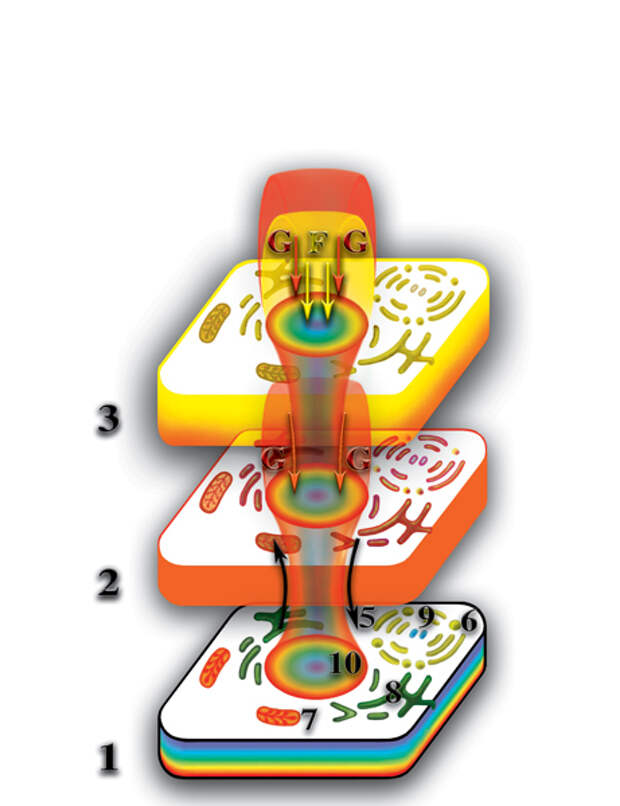
). Третьи материальные тела клеток многоклеточного организма также образуют жёсткую систему — третье материальное тело многоклеточного организма.
Появление третьих материальных тел у живых организмов явилось колоссальным качественным скачком в развитии живой природы. Наличие у клеток трёх взаимодействующих между собой уровней создало необходимые и достаточные условия для возникновения памяти, эмоций и интеллекта, что и является основой высокоорганизованной живой материи. Некоторые типы клеток многоклеточных организмов, при своей адаптации к выполняемым ими функциям, изменились до такой степени, что вызываемая ими деформация микропространства достигла четвёртого материального уровня планеты. Это — клетки головного, спинного и костного мозга. Аналогично, на этом уровне образуется четвёртое материальное тело многоклеточного организма из материальных тел клеток этого организма (Рис. 4.3.17
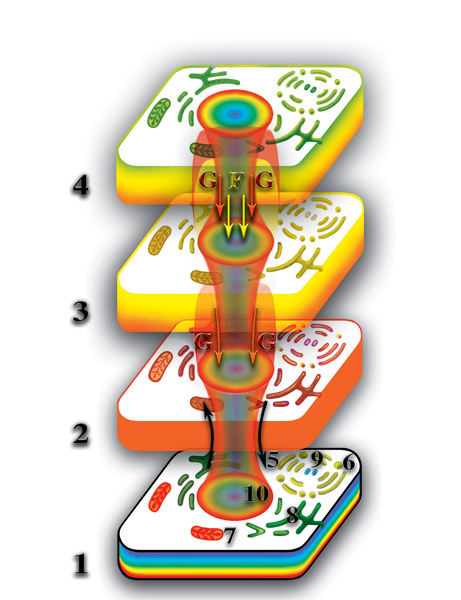{kind=link}
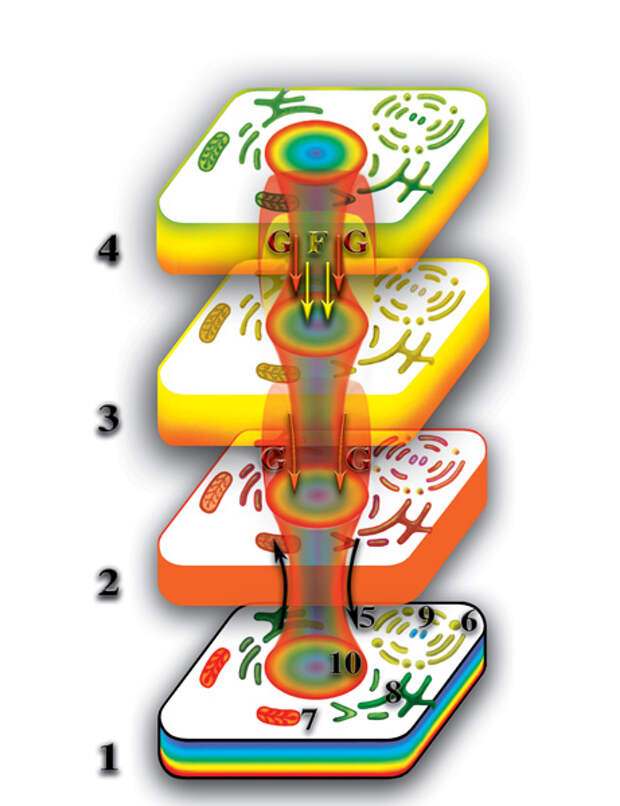
). Таким образом, в создании второго материального тела учавствуют все клетки физически плотного организма. В создании третьего материального тела — большинство клеток. Четвёртые материальные тела могут возникнуть лишь у некоторых видов живых организмов и то на определённом уровне их развития. В создании пятого материального тела принимает участие только часть клеток многоклеточного организма. Поэтому, четвёртое и пятое материальные тела качественно (внешне — тоже) отличаются, как от третьего, так и от второго материальных тел многоклеточного организма.
Физически плотное или первое материальное тело клетки, вместе со вторым, третьим, четвёртым и т.д. материальными телами, представляют собой одну систему — живой организм, живую материю. Только все вместе они создают чудо природы — живую материю, жизнь, эволюция которой закономерно приводит к зарождению разума — самосознания живой материи. При нарушении взаимодействия физически плотного тела клетки (первого материального тела) с остальными материальными телами, происходит нарушение функционирования и самой клетки. Прекращение циркуляции первичных материй между уровнями клетки приводит к смерти последней. Живая клетка не может функционировать без обратной связи со своими другими материальными телами. Так же, как и другие материальные тела клетки не в состоянии функционировать, без постоянного процесса распада молекул в физически плотной клетке. После остановки жизненных процессов, физически плотная клетка распадается на органические и неорганические молекулы. Этот факт не требует пояснений. Но, что происходит, при этом, с остальными материальными телами клетки? Разрушаются ли они наподобие физически плотного тела клетки или наблюдаются иные процессы и, если да, то какие?
Действительно, второе и т.д., материальные тела возникают в результате деформации микропространства, создаваемой физически плотной клеткой. Поэтому, первое что может прийти на ум, так это то, что и все остальные материальные тела клетки исчезают, при распаде физически плотного. Но, так ли это, вот, в чём вопрос?
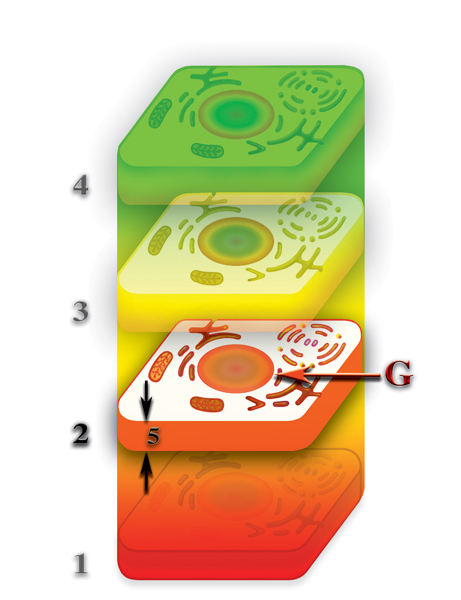
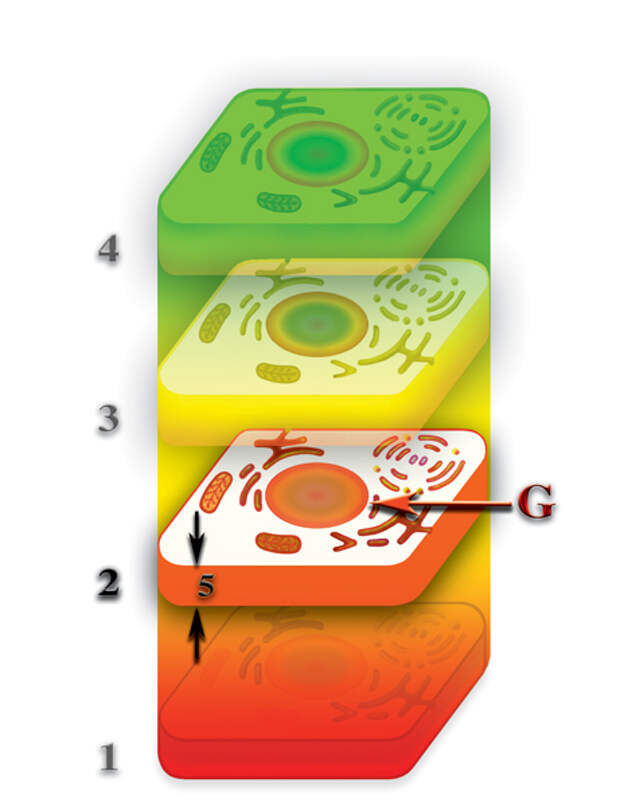
Давайте вспомним, что второе и т.д., материальные тела клетки появляются, как результат насыщения деформации, создаваемой клеткой на том или ином уровне первичными материями, которые не входят в состав каждого из этих уровней. Второе материальное тело — первичной материей G, третье — G и F, четвёртое — G, F и E и т.д. При таком раскладе, бросается в глаза качественное отличие первого от всех остальных материальных тел клетки. Первое материальное тело образовано из гибридной материи, возникшей при слиянии семи первичных материй. Все остальные материальные тела клетки возникли, как результат насыщения первичными материями деформации микропространства, создаваемой физически плотным телом клетки. Как след на мягкой почве заполняется дождевой водой, так и деформация микропространства, вызываемая физически плотной клеткой, насыщается соответствующими первичными материями. И, как не всегда исчезает след, оставленный ногой в мягкой почве, так и не всегда исчезают второе и другие материальные тела, после разрушения физически плотного тела клетки.
Давайте выясним, что же с ними происходит? При наличии у клетки только второго материального тела из первичной материи G, возникает ситуация в которой возможно несколько вариантов развития процесса. Второе материальное тело со временем теряет плотность насыщения первичной материей G; при наличии физически плотного тела, восполнение потерь происходит за счёт насыщения первичными материями, высвобождаемыми при распаде молекул внутри клетки, но и большая часть потерь вторым материальным телом первичной материи G, в то же самое время, вызвано возвратным потоком этой первичной материи на физически плотный уровень. Этот обратный поток и является необходимым условием нормального функционирования живой клетки. При разрушении физически плотной клетки, прекращается и обратный поток первичной материи G от второго материального тела к первому (Рис. 4.3.18
{kind=link}
).
Второе материальное тело продолжает терять некоторую часть первичной материи G, из которой и образуется собственно второе материальное тело клетки. В результате чего, плотность второго материального тела уменьшается, оно, как бы «тает». И если бы процесс «таянья» продолжался бы, безусловно, второе материальное тело клетки, через некоторое время после гибели физически плотного, исчезло бы. Но, этого не происходит. И вот, почему. Вспомним, что после завершения формирования планеты, первичные материи продолжают пронизывать зону неоднородности пространства, в которой произошёл синтез планеты. А это означает, что потоки первичных материй пронизывают и все материальные тела клетки, включая и физически плотное. И если насыщение физически плотного тела свободными первичными материями не играет принципиальной роли в функционировании физически плотной клетки, то, при пронизывании потоками первичных материй второго и других материальных тел клетки, картина кардинально меняется. Второе материальное тело представляет собой сгусток первичной материи G, заполнившей деформацию микропространства, созданную физически плотным телом на второй материальной сфере. Поэтому, при пронизывании первичными материями планетарного пространства, первичная материя G насыщает и второе материальное тело. Аналогично тому, как потеря воды лужей или водоёмом в жаркие дни возмещается посредством дождей. Главное — чтобы «дожди» шли регулярно. И если в случае с лужами это случается далеко не всегда, то в случае насыщения второго материального тела первичной материей G, подобная проблема практически никогда не возникает. Таким образом, качественное отличие природы образования физически плотного тела клетки от природы образования других материальных тел клетки, создают уникальную ситуацию, без которой эволюция живой материи была бы просто невозможной.
После разрушения физически плотной клетки, другие материальные тела клетки не исчезают, не разрушаются, а сохраняются за счёт подпитки потоками первичных материй, пронизывающих планетарное пространство.
Правда, существует весьма существенное отличие между этими двумя состояниями. Без физически плотного тела клетки, в котором происходит активный процесс распада молекул на первичные материи их образующие и мощного насыщения ими второго и других материальных тел клетки, вторичное насыщение первичными материями этих тел происходит очень медленно. В результате этого все процессы, происходящие на уровне второго и других, материальных тел клетки замедляются в сотни, а порой, и в тысячи раз. Замедляются, но не прекращаются. Это — весьма важный момент, имеющий принципиальное значение для понимания, как самой жизни, так и возможности эволюции живой природы.
Давайте подробно, ступенька за ступенькой, шаг за шагом, проанализируем происходящие процессы в живой системе после разрушения физически плотного тела клетки. При наличии у клетки только второго материального тела, после разрушения физически плотного тела клетки, второе материальное тело не исчезает, не рассеивается, как утренний туман, под лучами Солнца. Конечно, плотность второго материального тела без физически плотного значительно падает, но подпитка за счёт первичных материй, пронизывающих планетарные уровни не позволяет полностью «высохнуть» второму материальному телу. Почему это принципиально важно? Что случилось бы, если бы второе материальное тело клетки «высыхало» после разрушения физически плотного тела? Ничего «особенного», только то, что эволюции живой материи, появления разума «просто» не произошло бы. Вполне возможны ситуации, когда второе материальное тело может быть полностью разрушено в силу тех или иных причин, как, например, воздействие мощных вихревых потоков первичных материй, протекающих через планетарные уровни. Но, такие явления случаются не так часто и не создают глобальных проблем, не угрожают живой материи и её эволюции в целом. Но, вопрос, почему «невысыхание» второго и других материальных тел клетки, после разрушения физического тела клетки, является ключевым моментом для возможности эволюции живой материи и зарождения разума, отложим на некоторое время и вернёмся к качественным процессам, происходящим с клетками, имеющими разную качественную структуру.
Если живая клетка имеет, как второе, так и третье материальное тело, то, при разрушении физически плотного тела, без подпитки первичными материями, через расщепление молекул в физически плотной клетке, оказываются уже два материальных тела — второе и третье материальные тела. И, естественно, после прекращения «подачи наверх» физически плотной клеткой первичных материй «худеет», как второе, так третье материальные тела клетки. Но, опять-таки, не происходит исчезновение этих материальных тел после разрушения физически плотной клетки за счёт всё того же насыщения последних первичными материями, постоянно пронизывающими планетарные уровни. Отличие заключается в том, что третье материальное тело клетки подпитывается уже двумя первичными материями G и F. Причём, скорость насыщения первичными материями третьего материального тела клетки больше скорости насыщения второго материального тела, по одной простой причине. Потоки первичных материй, попадая в планетарную зону деформации, при своём движении через неё, вынуждены «просачиваться» через качественные барьеры шести планетарных материальных сфер. В результате этого скорость их движения замедляется. и скорость движения первичных материй, достигших второй планетарной материальной сферы, становится минимальной, по отношению к скоростям на всех других планетарных уровнях.
Кроме этого, планетарные качественные барьеры влияют на разные первичные материи неодинаково, в результате чего, происходит изменение соотношения первичных материй в общем потоке, и скорости их движения друг относительно друга всё больше и больше начинают различаться после прохождения каждого качественного барьера планеты. Что, в свою очередь, в значительной степени сказывается на соотношении между первичными материями на каждом планетарном уровне и, соответственно, на интенсивности процессов, протекающих на каждом из планетарных уровней. Поэтому, при наличии у клетки второго и третьего материальных тел, насыщение первичными материями G и F третьего материального тела клетки будет происходить быстрее, чем происходящее в тоже время насыщение первичной материей G второго материального тела. И, если принять во внимание, что «таяние» или другими словами потеря вторым и третьим материальным телами клетки первичных материй происходит, примерно, одинаково, то, в результате различной плотности и скорости движения первичных материй через второй и третий планетарные уровни, скорость насыщения первичными материями этих тел будет различная. Вследствие чего, насыщение первичными материями третьего материального тела будет происходить относительно быстрее, чем насыщение второго.
Конечно, это насыщение не идёт ни в какое сравнение с насыщением первичными материями этих тел, при наличии физически плотного тела клетки, но, тем не менее, в результате этого насыщения, возникает некий избыток, относительно второго материального тела, первичных материй в третьем материальном теле клетки. Относительный избыток концентрации первичных материй на уровне третьего материального тела, по отношению к второму материальному телу, приводит к тому, что между третьим и вторым материальными телами клеток возникает очень слабая циркуляция первичной материи G (Рис. 4.3.19
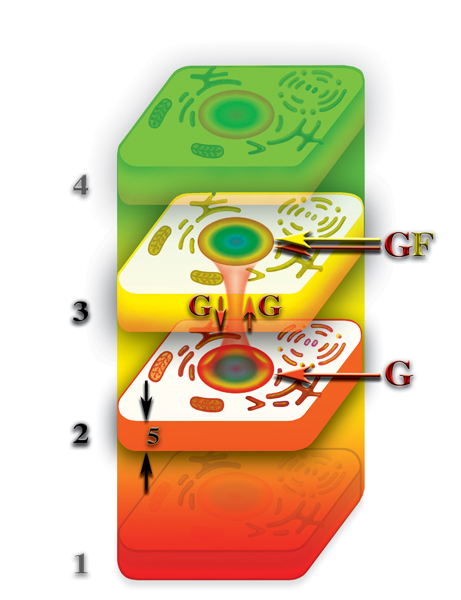{kind=link}
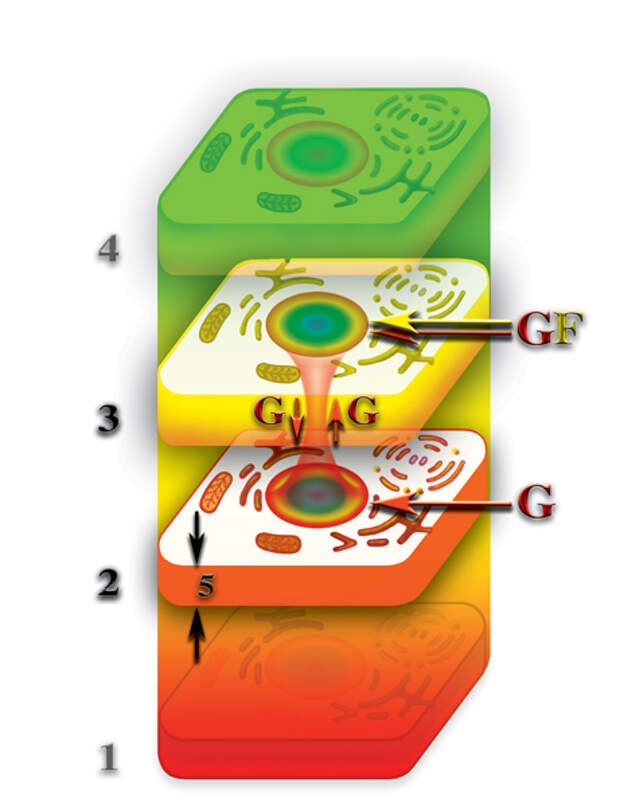
). Циркуляция первичных материй между вторым и третьим материальными телами после полного разрушения физически плотной клетки, есть ни что иное, как проявление жизнедеятельности. Другими словами, если физически плотная клетка перед тем, как разрушиться, имела в своей качественной структуре, как второе, так и третье материальные тела, то, после разрушения физически плотного тела, процессы жизнедеятельности клетки на этих уровнях не останавливаются, а только во много раз замедляются.
Аналогичные процессы происходят и на уровне физически плотных тел у земноводных и пресмыкающихся, когда при охлаждении тела активность их жизненных процессов замедляется в десятки раз без какого-либо вреда для этих животных. Более того, некоторые земноводные такие, как лягушки, могут полностью замёрзнуть, превратившись в ледяные статуи, а потом, нагреваясь под воздействием солнечных лучей, постепенно возвращаются к нормальному уровню активности. В таком замороженном состоянии они могут находиться сотни лет, только на те же сотни лет останавливается и развитие их организма, эволюция. В таком замороженном состоянии жизненные процессы физически плотных клеток организма лягушек замедляются в десятки тысяч раз, но не останавливаются полностью. Поэтому в замороженном состоянии лягушка продолжает использовать запасы органических молекул, накопленные в её клетках до того, как она замёрзла. Поэтому, в замороженном состоянии, лягушка очень медленно худеет, постепенно начинается голодание и, если вовремя не разморозить такую лягушку, она просто погибнет от истощения. Так как, к сожалению, в замороженном состоянии лягушки не в состоянии питаться. Мошкару, которая является основной пищей этих земноводных они пока ещё в состоянии ловить только в активном состоянии. Особой опасности умереть от истощения в замороженном состоянии у лягушек практически никогда не возникает, так как замораживаются они только на зимний период, когда температура среды их обитания опускается ниже нуля по Цельсию. Так вот, без физически плотного тела клетки во втором и третьем материальных телах жизненные процессы не останавливаются, а замедляются в тысячи раз. Но, тем не менее, это всё равно не полная смерть, которая подразумевает полную остановку жизненных процессов на всех уровнях, так называемую, абсолютную смерть.
Так вот, для живых организмов, в большинстве случаев, абсолютная смерть не наступает никогда.
После разрушения физически плотного тела клетки наступает относительная смерть живого организма, когда протекающие процессы жизнедеятельности на уровне второго и третьего материальных тел протекают в сотни тысяч раз медленнее, чем, при наличии плотного физического тела. При этом, живой организм теряет физически плотное тело, в данном случае одной клетки, но «верхние этажи» — второе и третье материальные тела — продолжают свою жизнедеятельность, хотя и замедленную в сотни тысяч раз. Правда, при этом, происходит эволюционное «замораживание» этих тел. К счастью, в таком состоянии эти тела не находятся вечно. Для одноклеточных организмов полное разрушение физически плотного тела происходит, при процессе деления (Рис. 4.3.20
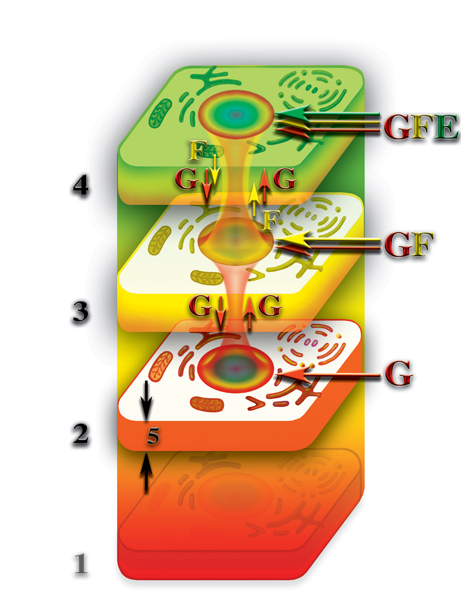{kind=link}
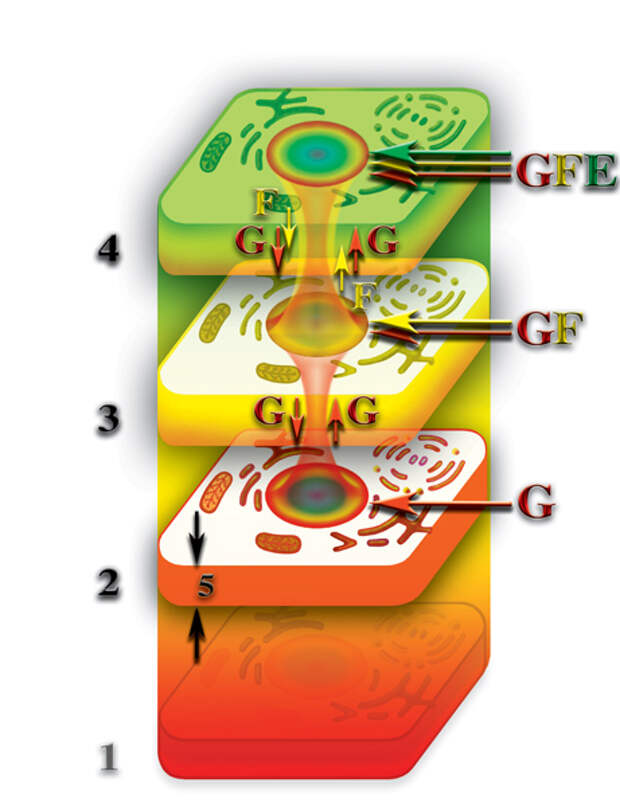
). В результате деления появляются две тождественные новые клетки, в то время, как «старая» клетка исчезает полностью разрушаясь во время процесса деления. Поэтому, «старая» клетка умирает, в том понимании, что она перестаёт существовать.
"
Это небольшая выдержка из одной из книг Николая Левашова ("Неоднородная Вселенная"). Если хотите узнать более подробно и не только про это, то все необходимые материалы имеются на сайте http://www.levashov.info
Продолжение следует....
Свежие комментарии